![]()
![]()
Katrai dzîvajai radîbai, kas
eksistį uz ðîs zemes, vajadzįja iziet grûtu konkurences ceïu, kâ rezultâtâ
organismi izveidoja daþâdus mehânismus, kas palîdzįja viōiem izkonkurįt
citus organismus, iekarojot jaunas niðas un iegûstot resursus savai eksistencei.
Atkarîbâ no organisma komplicįtîbas bija iespįjami daþâdi pielâgojumi.
Sâkotnįji tie bija daþâdu veidu toksîni, kurus organismi sekretįja apkârtįjâ
vidį, lai ar tiem nomâktu citus organismus. Izveidojoties nervu sistįmai,pavįrâs
principiâli jaunas iespįjas komplicįtai uzvedîbai. Kâ paðsaglabâðanâs un
aizsardzîbas mehânisms radâs aizsarguzvedîba. Ðajâ ziōâ interesantas ir
bites, to sociâlâ organizâcija ir relatîvi augsta. Uzmanîbas vįrta ir arî
biðu aizsarguzvedîba. Bieþa cilvįku saskare ar ðiem kukaiōiem ir rosinâjusi
par to aizdomâties. Jau senâs civilizâcijas zinâja par to, ka bites dzįlieniem
ir ârstniecisks efekts. Tâs izmantoja bites savâ medicînâ (reimatisku sâpju
ârstįðanâ). Bieþi vien ðie labvįlîgie kukaiōi var nodarît arî ievįrojamu
ïaunumu veselîbai.
Biðu indes aparâts ir evolûcijas
procesâ izveidots speciâli, lai nodroðinâtu biðu kolonijas aizsardzîbu
pret citiem kukaiōiem un zîdîtâjiem. Biðu indei ðajâ gadîjumâ ir izdzîvoðanas
loma, jo medus (biðu produkti) ir ïoti laba pârtika citiem kukaiōiem un
arî mugurkaulniekiem. Bitįm agresîva uzvedîba parasti nav raksturîga,
tâ parâdâs tikai biðu stropu tuvumâ. Kâ izpįtîja (Gary, 1974) smarþa ir
faktors, kas arî izraisa ðâdu agresîvu uzvedîbu. Pirmie zinâtniskie pįtîjumi
par biðu indi datįjami ar 19. gs. (Langer, 1897). Ðie primitîvie
neveiklie pįtîjumi jau tanî laikâ parâdîja ka biðu inde nav noteikta viela,
bet ka tâ sastâv no vairâku vielu maisîjuma. Daudz pilnîgâki rezultâti
tika iegûti tikai mûsu gadsimta 30-tajos gados, kad Neumanns ar lîdzstrâdniekiem
atklâja, ka biðu inde ir bioloėiski aktîvu vielu maisîjums. Jaunu biðu
indes ievâkðanas metoþu izstrâde pavįra iespįju ievâkt indes daudzumu,
kas ir pietiekams analîtiskâm metodįm. Tas savukârt veicinâja indes izpįti.
Izpįtes sâkumâ zinâtnieki saskârâs ar daþâdâm problįmâm; viena no tâdâm
bija tas, ka indes sastâvs daþâdos pįtîjumos bija atðíirîgs. Tas bija saistîts
ar to, ka biðu indes koncentrâcija variį atkarîbâ no sezonas un kukaiōa
vecuma, kâ arî bites statusa (darba bite, vai biðu mâte). Tâ, piemįram,
lielâkais indes daudzums ir biðu mâtįm, tas dod iespįju tâm iegût augstâko
varu hierarhijâ. Darba bites producį indi relatîvi mazâkos daudzumos; bitei
novecojot, sintįzes intensitâte strauji samazinâs.
Galvenâs grûtîbas biðu indes
pįtîjumos bija efektîvu un tîru metoþu trûkums biðu indes ievâkðanâ. Vienas
no pirmajâm indes ievâkðanas metodįm bija ïoti netîras un ar letâlu iznâkumu
kukainim. Ïoti mazus indes daudzumus varįja iegût, veicot biðu indes aparâta
sekciju (ðíįrsgriezumu), pįc kuras indi ekstrahįja no indes aparâta izvilkuma.
Vįlâk radâs otra jau labâka metode, tomįr ar nepietiekamu indes ieguves
daudzumu. Saskaōâ ar ðo metodi mehâniski stimulįja bites vįderiōu (to spaidîja),
tâ rezultâtâ kukainis izbîdîja dzeloni, cenðoties iedzelt. No ðî dzeloōa
arî savâca indes pilîtes. Mûsdienâs biðu indes iegûðanai izmanto efektîvâku
un humânâku metodi. Pįc ðîs metodes biðu indi var iegût gan laboratorijas,
gan lauka apstâkïos. Metodes bûtîba ir bites viegla stimulįðana ar augstfrekvences
elektrisko strâvu. Ja inde tiek iegûta laboratorijas apstâkïos, tad bites
ievieto speciâlâ plastmasas kamerâ, caur kuru impulsveidîgi laiþ cauri
augstfrekvences strâvu. Pįc pâris tâdâm procedûrâm kamera tiek atvįrta
un bites izlido, atstâjot uz kameras sieniōâm mazus indes pilieniōus, kurus
vįlâk vakuumâ þâvį un iegûto indes plįvîti noōem no sieniōâm. Atkârtojot
ðâdu procedûru neskaitâmas reizes, var iegût lielu diezgan tîras biðu indes
daudzumu. Metode, kuru izmanto lauka apstâkïos, nav tik tîra, tomįr ir
pietiekami efektîva un vienkârða. Biðu indi ievâc no speciâli sagatavotas
stropa skrejas, kas tiek stimulįta ar strâvu. Ikreiz, kad bite ielido vai
izlido no stropa, tâ ir pakïauta strâvas stimulam, kas momentâni izraisa
indes pilîtes izdalîðanos. Vįlâk inde no skrejas tiek noōemta un íîmiski
attîrîta no piemaisîjumiem.
Svaigi iegûta biðu inde ir
caurspîdîga, bezkrâsaina viela ar asu, nedaudz rûgtenu smarþu un skâbu
reakciju (pH <7).
Viegli ðíîst ûdenî, vai vâjâs
skâbįs. Sausa biðu inde ir pulverveida viela baltâ vai gaiði dzeltenâ
krâsâ. Pârdoðanâ esoðiem produktiem krâsa var mainîties no tumði pelįkas
lîdz brûnai. Krâsas atðíirîbas rodas uzglabâðanas rezultâtâ, fotooksidįjoties
bioloėiski aktîvâm vielâm (histidînam, triptofânam, fosfolipâzei A2
un melittînam).
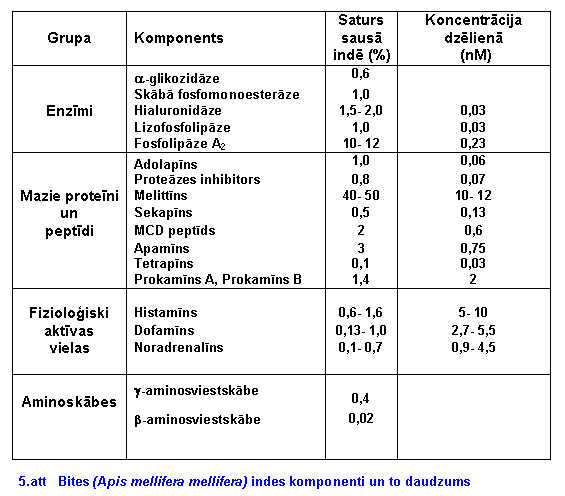
Indes sadalîðana pa sastâvdaïâm ir grûts un darbietilpîgs process. Lai to veiktu, izmanto daudz un daþâdas metodes, tâs kombinįjot (daudzpakâpju metode). Daudzpakâpju metode ietver sevî dialîzi zem spiediena, filtrâciju caur gįlu, jonu apmaiōas hromatogrâfiju. Tâlâko atdalîðanu veica balsoties uz analizįjamâ komponenta îpaðîbâm. Mûsdienâs vismaz 40 komponenti biðu indį ir izdalîti un noteikti.
a-Glikozidâze
(invertâze) E.C.3.2.1.20
Ðis enzîms tika atrasts gįla
filtrâcijas izejoðajâ frakcijâ, kopâ ar skâbo fosfomonoesterâzi. Abi enzîmi
ir skâbie glikoproteîni ar pH 0,5 ą 0,1.
a-glikozidâzes
daudzums sausâ indį ir aptuveni 0,6 % no masas. Molekulmasa tîrâ indį ir
170000. Maksimâlâ enzîma aktivitâte ar substrâtu p-nitofenil-a-glikopiranozîdu
tika novįrota pie pH 4.8 un 46°C. a-glikozidâze
ir netoksiska viela, spįcîgs antigįns, kas izraisa daudz antivielas truðiem.
Tâs termolabilitâti var izmantot kâ râdîtâju pareizai indes uzglabâðanai.
Ðis enzîms ir kopâ ar enzîmiem, kas transformį savâkto nektâru par medu.
Ðî enzîma klâtbûtne ir kâ râdîtâjs, ka dotâ inde tika iegûta ar elektriskâs
stimulâcijas metodi.
Skâbâ fosfomonoesterâze
(E.C.3.1.3.2)
Enzîms sausâ indį sastâda
1% no visas masas. Minimâlâ molekulmasa 55000. Lielâkâ aktivitâte
ar substrâtu p-nitrofenilfosfâtu novįrojama pie pH 4,7 un temperatûras
35°C. Enzîms termolabils, netoksisks ar spįcîgu antigenitâti (spįcîgs antigįns).
Ir pamats domât, ka ðî viela ir identiska Hoffmana izolįtajai vielai ko
nosauca par alergįnu B. Nav arî zinâmu gadîjumu, kad kâds ir saindįjies
ar ðo enzîmu. Turklât pįc (OConnor and Peck, 1980) ðis enzîms, tâpat kâ
arî a-glikozidâze, nav atrodams ultratîrâ biðu indį. Iespįjams, ka pįc
stimulâcija metodes, ko izmanto komerciâlie raþotâji, enzîms varįtu rasties
atmiruðo sekretoro ðûnu lîzes rezultâtâ.
Hialuronidâze (E.C.4.2.2.1)
Tâ izsauc starpðûnu telpâ esoðo
viskozo mukopolisaharîdu, hidrolîtisku depolimerizâciju. Tâdâ veidâ tâ
veicina injicįtâs indes izplatîðanos apkârtįjos audos. Vielas klâtbûtni
biðu indį atklâja 1954 gadâ Neumanns un Habermanns. Seviðíu interesi zinâtnieki
veltîja ðai vielai, jo saistîja tâs iespįjamo sakaru ar anafilaktisko reakciju
uz bites dzįlienu. Tâ ir viela ar molmasu 40000. Stabila pret daþâdâm fizikâlâm
iedarbîbâm, (saldįðana, þâvįðana un uzglabâðana). Tika konstatįts, ka tiem
pacientiem, kam ir alerėija pret biðu indi, ir paaugstinâts IgE uz hialuronidâzi.
Tas varįtu bût alergįns ðiem pacientiem. Tâs daudzums indes pûslîtî pieaug
daþu dienu laikâ, bitei sasniedzot noteiktu vecumu, un tâds arî saglabâjas
visâ atlikuðajâ dzîves laikâ. Nav arî novįrojamas nekâdas sezonâlas
fluktuâcijas. Atðíirîbas ir bites tipâ, piemįram, parastajai darba bitei
ðîs vielas daudzums indį ir 5 reizes lielâks nekâ biðu mâtei. Tas, domâjams,
ir izskaidrojams ar konkurenci, kas ir starp ðîm bitįm. Biðu mâte dzeï
tikai otru biðu mâti, lai iegûtu augstâku varu. Savukârt darba bitįm nepiecieðams
aizsargâties pat no zîdîtâjiem.
Lizofosfolipâze (E.C.3.1.1.5)
Doerijs un Pearsons 1964. gadâ
atklâja nelielu fosfolipâzes B aktivitâti biðu indį. Vįlâk (1982. gadâ)
ar to strâdâja Ivanova un Ðekenderjovs. Viōi no biðu indes izdalîja vielu
ar molmasu aptuveni 22000 ą 2000. Optimâlâkie apstâkïi enzîma darbîbai
ir pie pH 9. Ðî viela veicina intoksikâcijas procesu, jo enzîms regulį
lizolecitîna lîmeni (neïauj tam pieaugt). Lizolecitîns inhibį fosfolipâzes
A2 aktivitâti. Tâ rezultâtâ fosfolipâzes inhibitors tiek nomâkts
un ferments var strâdât. Arî papildu taukskâbju producįðana pastiprina
lîtisko procesu. Ðajâ piemįrâ var labi redzįt dzîvnieku indes komponentu
pareizo balansu.
Fosfolipâze A2
(E.C.3.1.1.4)
Tâ pieder pie enzîmu grupas,
kas hidrolizį dabiskos lipîdus. A2 veids katalizį daþu lipîdus
deacilâciju pozîcijâ 2 , tâdâ veidâ veidojot lizofosfoglicerîdus un garâs
taukskâbju íįdes. Abi produkti ir stipri "membrânaktîvie aėenti". Ja fosfolipîdi
ir hidrolizįti, membrânas funkcionâlâ integritâte var bût izjaukta, un
ðûnas aiziet bojâ. Ðo mehânismu ir izmantojuði savâs indįs arî citi dzîvnnieki.
Skatoties no otras puses, ðis enzîms ir organismâ visur, ðim enzîmam ir
intracelulârâ regulatora nozîme. Fosfolipâze A2 ir iesaistîta
prostaglandînu un leikotriįnu metabolismâ, glicerolipîdu metabolismâ un
fosfolipîdu degradâcijâ. Fosfolipâzes A2 klâtbûtni biðu indį
pirmo reizi konstatįja Neumanns un Habermans 1954. gadâ. Enzîma izolįðanu
mįėinâja vairâkas zinâtnieku grupas (Habermann and Neumann; Munjal and
Elliot, Shipolini).

1.att. Fosfolipâze A2
Oa -Adolapîns
Bulgâru zinâtnieki Ðekenderjovs
un Koburova 1982. gadâ atklâja jaunu proteînu biðu indį. Tas bija izolįts
no Oa frakcijas (Habermanna nosaukta) divpakâpju izdalîðanâ. Viela ir vienkârðs
polipeptîds, kas sastâv no 103 aminoskâbju atlikumiem, iztrûkstot cisteînam,
metionînam un triptofânam, ar minimâlo aprįíinâto molmasu 11500. Glicîns
tika identificįts vienîgi kâ N gala atlikums. Attîrîts materiâls parâdîja
stipru inhibįjoðu iedarbîbu uz kaía liesas mikrosomâlo ciklooksigenâzi
un lipooksigenâzi. Abi augstâkminįtie enzîmi ir iesaistîti arahidonskâbes
transformâcijas pirmajos soïos prostaglandîna biosintįzį. Lielâkâ daïa
nesteroîdo pretiekaisuma lîdzekïu, kâ zinâms, inhibį endoperoksîdu metabolîtu
producįðanu. Pelįm ðî viela izraisîja stipru analgįtisku iedarbîbu (kontrolei
tika pielietoti Randall-Selittos testi). Citos eksperimentos tas uzrâdîja
arî antipirįtiskas îpaðîbas. Savu nosaukumu viela ieguva, pateicoties izcelsmei
un atklâðanas vįsturei.
Proteâzes inhibitors.
Ðekenderjova grupa pirmo reizi
ziōoja, ka biðu indei piemît izteikta inhibįtspįja uz tripsîna proteolîtisko
aktivitâti. No daþâdiem avotiem ievâktâ biðu inde tika pârbaudîta, un tika
konstatįta tâda pati aktivitâte. Pįc sareþėîtas atdalîðanas metodes tika
izolįti divi proteîni, kas bija lîdzîgi pįc savâm îpaðîbâm. To kopįjais
daudzums no biðu indes masas bija aptuveni 0,8 %. Abi komponenti ir polipeptîdi,
kas sastâv no 63- 65 aminoskâbju atlikumiem. Proteîna sastâvâ nav treonîna,
metionîna un histidîna. Astoōi cisteîni ir savienoti ar 4 disulfîdsaitįm.
Neitrâlâ un skâbâ pH inhibitori ir termiski stabili. Vįlâk, turpinot pįtîjumus,
grupa pįtnieku no Londonas Universitâtes Koledþas izolįja divas ïoti lîdzîgas
olbaltumvielas H3 un H1. Vienîgâ atðíirîba, kas bija starp ðîm vielâm,
bija tas, ka peptîdam H1 iztrûka pirmie 4 atlikumi N galâ. Pįtîjumi parâdîja,
ka dabâ nav atrasta homoloėija ar citiem proteolîtiskajiem inhibitoriem.
Ir viedoklis par to, ka ðîs vielas funkcija kâ inhibitoram nav primâra.
Tâ kâ ðî viela nav toksiska cilvįkam, varįtu bût, ka tâ ietekmį kukaiōus.
Melittîns.
Galvenais biðu indes komponents
ir polipeptîds, kas sastâv no 26 aminoskâbįm un tâ molmasa ir 2840. Habermanns
nosauca ðo polipeptîdu par melittînu. Tâ klâtbûtni noteica Neumanns un
lîdzstrâdnieki (1952.), kâ tieðo hemolizînu FI. Mûsdienâs tas ir vislabâk
izpįtîtais komponents ne tikai biðu indį, bet arî no peptîdiem ar hemolîtiskâm
îpaðîbâm. (Habermann and Jentisch, 1967.) noteica proteîna sekvenci. Vįlâk
sekvenci pârbaudîja, sintezįjot ðo proteînu Lubke ar lîdzstrâdniekiem (1982).
Terwilligers un Eisenbergs noskaidroja melittîna 3D struktûru, izmantojot
rentgeno-kristalogrâfiju. Tika konstatįts, ka dabâ ðî viela pastâv divos
veidos. Kâ pirmo var minįt stâvokli ûdens vidį- bites dzelonî. Faucons
ar kolįėiem atklâja , ka ûdens ðíîdumâ viela pastâv lîdzsvarâ: monomįrs
Ŧ oligomįrs -tetramįrs (2.att). Ðis lîdzsvars ir atkarîgs no fizikâlajiem
parametriem (temperatûras un jonu spįkiem). Otra vide, kurâ bieþi atrodas
melittîns, ir fizioloėiskie ðíîdumi, tajos tas veido monomįru. Koncentrâcijâ
lîdz 4 Ũ 10 -3 M (pįc Davsona).

2.att. Melittîna tetramįrs
Proteîns monomįra formâ spįj seviðíi labi saistîties ar fosfolipîdiem. Mijiedarbîbu un saistîðanos ar fosholipîdu slâni pastiprina paaugstinâta temperatûra, arî membrânas fluiditâte var pastiprinât mijiedarbîbu. Pazeminot temperatûru zem kritiskâs, saistîtais melittîns tiek atbrîvots. Lîdzîga îpaðîba piemît daudziem membrânas proteîniem, citohromam b5, un citohrom-b5-reduktâzei. Melittîna citotoksiskais efekts izpauþas kâ ðûnas membrânas dezintegrâcija, tâdâ veidâ padarot iespįjamu tâs hidrolîzi ar citiem peptîdiem , kâ arî izjaucot membrânas citas struktûras. Ir zinâms ka melittîns spįj inhibįt arî proteînkinâzes. Aptuvenâ melittîna koncentrâcija biðu indį ir 17-21 mM (kâ monomįrs). Vielas aptuvenais daudzums vienâ dzįlienâ ir 10-12 mM, Atðíaidîtam ar iekððûnas ðíîdumiem tâ koncentrâcija varįtu bût aptuveni 10 -4 M, pie kuras ir novįrojama arî hemolîze. Ðâds daudzums izsauc apkârtįjo ðûnu lîzi un veido lokalizįtu iekaisumu. Jebkurð no ðiem efektiem var tikt pastiprinâts ar fosfolipâzi A2. Vįlâk zinâtnieku grupa no Čikâgas Universitâtes (Degrado et. Al. 1981) sasaistîja polipeptîda uzbûvi ar tâ bioloėisko aktivitâti. Molekula sastâv no diviem domįniem. Viens domįns ir N-terminâls no 20 atlikumiem ar hidrofobo indeksu virs 2 ar tendenci uz a íįdes konfigurâciju ar ampifîlâm îpaðîbâm. Otrs ir C terminâla domįns ar aptuveni 6 atlikumiem, tas ir izteikti hidrofîls. Labâkai darbîbas mehânisma izpratnei, nolįma uzsintezįt lîdzîgu vielu, kurai N terminâla a íįde bûtu ar mazâko homoloėiju dabiskajam peptîdam. Sintezįtâ viela izrâdîjâs 2,5 reizes hemolîtiski aktîvâka par dabisko. Tâdejâdi zinâtnieki nonâca pie secinâjuma, ka homolîtiskâs îpaðîbas atspoguïojas N terminâla a íįdes hidrofobicitâtį. Peptîda augstâ membranolîtiskâ aktivitâte uzvedinâja uz domas, ka tas nevarįtu bût sintezįts indes aparâta celulârajâ daïâ, aktîvâ formâ. Tika noskaidrota gįna struktûra, kas satur informâciju par melittîna sintįzi. Tas tika izdalîts no indes dziedzera. Melittîna mRNS tika translįta un iegûts melittîna prekursors- propromelittîns. (Suchanek and Kreil, 1977). Lai pârvįrstu to galaproduktâ, ir nepiecieðamas vairâkas enzimâtiskâs reakcijas. Prefragments tiek izðíelts ar "signâla" tipa endopeptâzi, kas atrodas indes dziedzerî, pârðíeï vienkârðâs saites A21-A22, (Kaschnit and Kreil, 1978). Nâkoðais solis iekïauj C-terminâla glutaminil glicîna pįrvįrðanu par glutaminilamîdu. Ðîs amidâcijas mehânisms, iespįjams, ir kopîgs visiem biðu indes peptîdiem ar amidįtiem C-terminâliem. Prosekvences izðíelðana tiek nodroðinâta pakâpeniskâ reakcijâ ar dipeptidilpeptidâzi IV, kas ir indes dziedzerî. Enzîms nodala dipeptîdus kuri vįlâk ar dipeptidâzi tiek degradįti lîdz brîvâm aminoskâbįm (dipeptidâze arî ir indes dziedzerî). Ðâds mehânisms nodroðina to, ka pa tâdu laiku pįdįjais aktivâcijas solis tiek pabeigts tâdâ indes aparâta daïâ, kur tam nav kontakta ar ðûnu struktûrâm, kas satur fosfolipîdus (piemįram bites indes dzelonis, kas sastâv no hitîna).
Sekapîns
Tas ir biðu indes komponents,
kas indį ir mazos daudzumos (mazâk nekâ 0,5% no sausâ produkta). Sekapîna
struktûras noteikðanu un izolįðanu veica 1976.gadâ (Gauldie et al). Tâ
struktûra nelîdzinâs ne viena biðu indį esoðâ peptîda struktûrai. Peptîds
praktiski ir netoksisks zîdîtâjiem (pelįm LD50 40mg/kg). Kreils
atklâja ðî proteîna gįnu, kas kodį preprosekapînu. Peptîda koncentrâcija
biðu mâtes indes pûslîtî ir lielâka nekâ parastu darba biðu pûslîðos. Iespįjams,
ka sekapînam ir kâda iedarbîba uz kukaiōiem, vai arî tas aktivį citus biðu
indes komponentus.
MCD (401 peptîds)
Pirmo reizi to konstatįja Fredholms
1966. gadâ. Atðíirîbâ no citâm indį esoðajâm vielâm, (melittîna un fosfolipâzes),
tas spįj izsaukt histamîna izdalîðanos no þurkas vįderplįves tuklajâm ðûnâm.
Tâ primârâ struktûra (3.att.) tika noskaidrota neatkarîgi un gandrîz
tajâ paðâ laikâ abâm zinâtnieku grupâm (Haux, 1969; Vernon et al., 1969).
Peptîda molekulmasa ir aptuveni 5000-6000. Tas ir termostabils peptîds,
to stabilizį galvenokârt divas disulfîdsaites.

3.att. MCD peptîda struktûrformula
MCD toksicitâte intraventrikulâri
pelįm LD50 20mg/1 kg. Pįc vielas ievadîðanas dzîvniekiem tika
novįrota nekoordinįta hiperaktivitâte, kam sekoja elpoðanas paralîze (Habermann
1977). Eksperimentos uz þurkâm, tâs tika injicįtas ar 5 mg/kg lielu MCD
devu. Tâ rezultâtâ dzîvnieki aizgâja bojâ no histamîna intoksikâcijas.
401 peptîdam nosaukumu deva Habermanns, balstoties uz tâ fizioloėisko efektu
(Mast Cell Degranullating peptide). Peptîds izraisa iekaisumu. Vietâ, kur
tas ir injicįts, lokâli izdalâs histamîns un serotonîns. Ir zinâms, ka
MCD var degranulįt þurkas vįderplįves tuklâs ðûnas bez lîzes koncentrâcijâ
10- 9 M. Iepriekðįjie pįtîjumi râdîja, ka MCD peptîds spįj konkurįt pįc
dendrotoksîna saistîðanâs vietâm, kas ir saistîts ar ar potenciâla sensatîviem
K+ kanâliem. Tâdejâdi MCD var strâdât kâ katjonu kanâlu antagonists.
Tetrapîns
Tetrapîns biðu indį ir
ïoti mazos daudzumos (mazâk kâ 0,1% no sausâs indes masas). Dzįliens satur
aptuveni 70 mg. Tâ izolįðanu no biðu indes pirmo reizi 1976. gadâ veica
Gauldie, un primâro struktûru noskaidroja Ovčinōikovs 1980.gadâ. Tetrapîna
struktûra ir lîdzîga apamîna un MCD peptîda struktûrai. Tetrapîna toksiskums
nav pârâk augsta, pelįm LD50 ir 40mg/kg. Tas degranulį tuklâs
ðûnas , bet tâ spįja ir tikai 20 % no MCD peptîda.
Apamîns
Ðî peptîda izolâciju pirmo
reizi veica Habermans un Reizs 1966.gadâ. Tâ struktûru atklâja reizį divas
zinâtnieku grupas (Haux et al., 1967; Shipolini et al., 1967). Disulfîdsaiðu
pozîcijas noskaidroja (Callewaert et. al.,1967). Apamîna îpaðîbas tika
pįtîtas, izmantojot izotoksînu sintįzi, kâ arî modificįjot dabîgo peptîdu.
Apamîns ir biðu indes galvenais toksîns (neirotoksîns). Pelįm ievadot perifįri,
saindįðanâs simptomi sâk parâdîties jau pįc pâris minûtįm ar ïoti raksturîgu
ainu, nekoordinįta hipermobilitâte lîdz pat 12 stundâm. Hiperuzbudinâmîba
saglabâjas lîdz 2-3 dienâm. Letâlas devas izsauc kloniskas konvulsijas
un elpoðanas paralîzi, kam seko nâve (Habermanns, 1972).

4.att. Apamîna telpiskâ struktûra
Pelįm LD 50 i. v.
ir 3,8 mg/kg. Inde darbojas ðíįrsojot hematoencifalisko barjeru un iedarbojoties
uz CNS. Apamînam piemît liela alergįnâ aktivitâte. Tâ maksimâlâ aktivitâte
ir pie pH 9. Banks ar lîdzstrâdniekiem pirmo reizi izpįtîja, ka apamîns
selektîvi bloíį Ca2+ atkarîgos K+ kanâlus. Vielas
molekulmasa ir 2027 .Tas labi ðíîst ûdenî un fizioloėiskâ ðíîdumâ.
Prokamîni
Tâ ir grupa no diviem, varbût
trim homologiem peptîdiem (Nelsona peptîds, prokamîns A un prokamîns B),
ar kopįju unikâlu îpatnîbu, viōiem visiem pie C terminâla ir pievienots
histamîns. To no biðu indes izolįja Nelsons un OConnors 1968. Tâ struktûru
noteica Pecks un OConnors, 1978. gadâ. Peptîdu bioloėiskâ aktivitâte ir
saistîta ar antiradioaktîvu efektu. Mâkslîgi sintezįtajam glicilhistamînam
ir lîdzîgas antioksidatîvas îpaðîbas (Peck et al., 1978). Domâjams, ðâdas
îpaðîbas nodroðina histamîna mijiedarbîba ar brîvajiem radikâïiem, kas
rodas no jonozįjoðâ starojuma organismâ.
Atseviðíi grupas pârstâvji ar
spįcîgâm farmakoloėiskâm îpaðîbâm ir atrasti biðu indį, kâ arî parasti
ir čûsku indįs.
Histamîns.
Ðis ir visbieþâk sastopamais
Hymenoptera biogįnais amîns. Histamîna lîmenis indį variį atkarîbâ no vecuma.
35- 45 dienas vecâm bitįm tas ir maksimâlâ lîmenî (1900 ą 860 ng uz indes
pûslîti). Tas ir aptuveni 17 ą 8 nM (Oven et al.,1977). Tâ toksiskums zîdîtâjiem
variį, bet pamatâ ir zema- pelįm LD50 1300 mg/kg. Domâjams ka
tâdâ daudzumâ, kâdâ tas ir vienâ dzįlienâ, (aptuveni 6,3 nM), tas nevarįtu
izsaukt nekâdu intoksikâciju. Tâ efekts varįtu bût asinsvadu caurlaidîbas
palielinâðanâs un to dilatâcija, kas varįtu izsaukt sâpes dzįliena vietâ.
Iespįjamâ histamîna nozîme biðu indį ir kateholamînu izdalîðana.
Divi ðîs grupas pârstâvji ir
atrasti biðu indį (dofamîns un noradrenalîns) (Owen, 1971; Bank et al.,1976)
Dofamîns
Tâ daudzums biðu indį ir atkarîgs
no bites vecuma. Jaunâm bitįm daudzums indes pûslîtî ir zems (20
ą 40 ng uz pûslîti).Tas pieaug 20-25 dienas vecâm bitįm (1500 ą 200 ng
uz pûslîti). Dofamîna daudzums variį arî atkarîbâ no sezonâlâm svârstîbâm,
maksimums ir(4307 ą 1138 ng uz indes aparâtu) augusta vidû (Owen un Bridges,
1982). Dzįlienâ esoðâ dofamîna daudzums svârstâs no 2,7-5,4 nM. Kukaiōu
fizioloėijâ ðim amînam ir nedaudz atðíirîga nozîme nekâ zîdîtâju fizioloėijâ.
Tas ir iesaistîts kutikulas veidoðanas un cietinâðanas procesâ. Tas ir
arî tuvâkais noradrenalîna priekðtecis. Dofamîns var bût arî kâ neiromediâtors
daþiem kukaiōiem. Ir uzskats, ka dofamîns indį darbojas, mijiedarbojoties
sinerėiski ar kâdâm membranoaktîvâm vielâm. Gadîjumos, kad biðu inde tiek
pielietota pret kukaiōiem, dofamîns lielos daudzumos var izraisît kukaiōa
NS paralîzi.
Noradrenalîns
Ðis amîns tiek uzskatîts par
neiromediâtoru visiem kukaiōiem. Tâpat kâ dofamînam, tâ koncentrâcija indį
variį atkarîbâ no kukaiōa vecuma un sezonas, maksimums ir 40 dienas vecai
bitei (1880 ą 170 ng uz indes pûslîti). Noradrenalîna daudzums bites dzįlienâ
ir aptuveni 5 nM. Tâds daudzums ir par maz, lai varįtu izraisît kâdu farmakoloėisku
efektu uz zîdîtâjiem, tomįr kukaiōiem tas var izraisît CNS traucįjumus.
Aminoskâbes
19 brîvâs aminoskâbes tika
atrastas biðu indį (Nelson & OConnor, 1968). Kopumâ tâs ir tikai mazâk
nekâ 1% no sausâs indes svara. Ir noskaidrots, ka tikai daþas ir kâ pastâvîgs
indes komponents, kamįr citas tikai piedalâs indes biosintįzį. Tikai g-aminosviestskâbe
(0,04%) un b-aminosviestskâbe (0,02%) ir indes sastâvâ.
Ogïhidrâti
Biðu indį tika atrasti
tikai divi cukuri- glikoze (0,7%) un fruktoze (0,9%) no sausâs indes masas.
Abi ir arî hemolimfâ, un kalpo kâ galvenais enerėijas avots bitįm.
Lipîdi
Biðu indį lipîdi ir kopumâ
lîdz 5 % no sausâs indes svara.
Bites indes iedarbîba uz cilvįka organismu variį individuâli. Indį (5.att) ir vairâki alergįni. Stiprâkie no tiem ir skâbâ fosfatâze, melittîns, hialuronidâze un fosfolipâze A2. Viens bites dzįliens jûtîgiem cilvįkiem var izsaukt alerėisku reakciju, kuras novįrðanai vajag sniegt pirmo medicînisko palîdzîbu. Retâki gadîjumi ir tad, kad cilvįku sadzeï daudz bites (vairâk kâ 50), tâdi gadîjumi beidzas letâli. Galvenâ problįma ðajâ gadîjumâ ir organisma âtra reakcija uz indi (galvenokârt apamîna neirotoksiskuma dįï). Praksį ir bijuði gadîjumi, kad pie masveida biðu kodieniem (vairâk nekâ 50) cilvįks aizgâja bojâ no sistįmiskâs intoksikâcijas. Ðinî gadîjumâ nâve var iestâties pįc 23-24 stundâm. Zinâtnieki vairâkus gadus strâdâja pie ðî jautâjuma, jo masveida biðu dzįlienu rezultâtâ ir novįrojama augsta letalitâte. Pįdįjo gadu pįtîjumi bija ar panâkumiem, un tika izveidotas un pârbaudîtas daþâdas vakcînas. Vienu no tâdâm 1999. gadâ izstrâdâja Britu zinâtnieku grupa (Jones RG, Corteling RL, Bhogal G, Landon J) no Íîmiskâs Pataloėijas departamenta, St. Bartalomeja hospitâlî. Viōu vakcîna spįj neitralizįt biðu indes iedarbîbu daþu stundu laikâ, sistįmiskâs intoksikâcijas gadîjumâ. Vakcînas daudzums, kas ir nepiecieðams indes neitralizâcijai, ir aptuveni 20 reizes vairâk nekâ saōemtâs indes daudzums. Vakcîna varįtu bût efektîva tikai pie masveida biðu dzįlieniem, nepiecieðamâs vakcînas daudzumu izrįíina, balstoties uz dzeloōu skaitu uz vienu organismu. Vakcîna tiek testįta un, iespįjams, jau drîz bûs pieejama mediíiem. Pįtîjumi ðajâ jomâ turpinâs un, iespįjams, jau drîz bûs jaunâki un daudz efektîvâki lîdzekïi pret biðu kâ arî citu indîgo kukaiōu dzįlieniem.
Anthony T. Tu.,. INSECT POISONS, ALLERGENS, AND OTHER INVERTEBRATE VENOMS. 1984. Marcel Dekker Inc.
Assem, E. S. K., and Atkinson, G. (1973). Histamine release by MCDP (401), a peptide from the venom of honey bee. Br. J. Pharmacol. 48:337-338.
Bachmayer, H., Kreil, G., and Suchanek, G. (1972) . Synthesis of prome- littin and melittin in the venom gland of queen and worker bees: Patterns observed during maturations. J. Insect Physiol. 18:1515-1522.
Banks, B. E. C., Brown, C., Burgess, G. M., Burnstock, G., Claret, M., Cocks, T. M., and Jenkinson, D. H. (1979b) . Apamin blocks certain neurotransmitter-induced increases in potassium permeability. Nature 282:415-417.
Banks, B. E. C., Dempsey, C., Pearce, F. L., Vernon, C. A., and Whol- ley, T. E. (1981). New method of isolating bee venom peptides. Anal. Biochem. 116: 48-52.
Banks, B. E. C., Dempsey, C., and Yamey, J. (1982). Anti-inflammatory activity in the venom of Apis mellifera. Toxicon 20:4.
Banks, B. E. C., Dempsey, G., Vernon, C. A., and Yamey, J. (1980). The The mast cell degranulating peptide from bee venom. J. Physiol. (Lond. ) 308: 95-96P.
Banks, B. E. C., Garman, A. J., and Habermann, E. (1978). Structure- activity studies on apamin and mast cell degranulating peptide (MCDP)- 401. J. Physiol. (Lond. ) 284: 160-161P.
Banks, B. E. C., Hanson, J. M., and Sinclair, N. M. (1976). The isola- tion and identification of noradrenaline and dopamine from the venom of the honey bee, Apis mellifica. Toxicon 14:117-125.
Banks, B. E. C., Sinclair, N. M., and Vernon, C. A. (1979a). The poly- peptide components of bee venom, Habermehl and Mebs (Eds. ) . Proc. 3d Symposium of Plant, Animal and Microbial Toxins, London, pp. 65-78.
Billingham, M. E. J., Morley, J., Hanson, J. M., Shipolini, R. A., and Vernon, C. A. (1973). An anti-inflammatory peptide from bee venom. Nature 245: 163-164.
Bradbury, A. F., Finnie, M. D. A., and Smyth, D. G. (1982). Meehanism of C-terminal amide formation by pituitary enzymes. Nature 298:686-688.
Dotimas EM, Hamid KR, Hider RC, Ragnarsson U, Isolation and structure analysis of bee venom mast cell degranulating peptide, in Biochim Biophys Acta 1987 Feb 25;911(3):285-93
Gauldie J, Hanson JM, Shipolini RA, Vernon CA., The structures of some peptides from bee venom, in Eur J Biochem 1978 Feb;83(2):405-10
Hider RC, Ragnarsson U., A comparative structural study of apamin and related bee venom peptides, in Biochim Biophys Acta 1981 Jan 30;667(1):197-208
Jones RG, Corteling RL, Bhogal G, Landon J., A novel Fab-based antivenom for the treatment of mass bee attacks, in Am J Trop Med Hyg 1999 Sep;61(3):361-6
Koburova KL, Michailova SG, Shkenderov SV., Further investigation on the antiinflammatory properties of adolapin--bee venom polypeptide, in Acta Physiol Pharmacol Bulg 1985;11(2):50-5
Sandberg, B. E. B., and Ragnarsson, U. (1978). Solid phase synthesis of apamine, the principal neurotoxin in bee venom. 1nt. J. Pept. Protein Res. 11:23S-245.
Schoch, P., and Sarpent, D. F. (1980). Quantitative analysis of the bind- ing of melittin to planar lipid bilayers allowing for the discretecharge effect. Biochim. Biophys. Acta 60Z: 234-247.
Schröder, F,, Lubke, K., Lehmann, M., and Beetz, I. (1971) . Haemolytic activity and action on the surface tension of aqueous solutions of synthetic melittin and their derivatives. Experientia 27: 764-766.
Sessa, G., Freer, J. H., Colacicco, G., and Weissman, G. (1969). Interaction of a 1ytic polypeptide, melittin, with lipid membrane systems. J. Biol. Chem. 244: 3575-3582.
Shier, W. T. (1977a). Inhibition of acyl coenzyme A; Lysolecithin acyl- transferases by local anesthetics, detergents, and inhibitors of cyclic nucleotides phosphodiesterases. Biochem. Biophys. Res. Commun. 75 : 186- 193.
Shier, W. T. (1977b). Inhibition of prostaplandin biosynthesis by lysoleci- thin. Biochem. Biophys. Res. Commun. 78:1168-1174.
Shier, W. T. (1979). Activation of hiph levels of. endogenous phospholipase A2 in cultured cells. Proc. Natl. Aead. Sci. USA 76:195-199.
Shier, W. T. (1980). Activation of self-destruction as mechanism of action for cytolytic toxins, in Natural Taxins, Proc. 6th International Sympo- sium of Animla and Mierobial Taxins, Uppsala, August 1979, D. Eaker and I. Nadstrom (Eds.), Pergamon Press pp. 193-200.
Shkenderov S, Koburova K., Adolapin--a newly isolated analgetic and anti-inflammatory polypeptide from bee venom, in Toxicon 1982;20(1):317-21
Terwillinger, T. C. and Eisenberg, D., The structure of melittin, I. Structure determination and partial refinenment, in "J. Biol. Chem.", Vol.257,pp. 6010-6015, 1982
Terwillinger, T. C. and Eisenberg, D., The structure of melittin, II. Interpretation of the structure in "J. Biol. Chem.", Vol.257, pp.6016-6021, 1982
Vetter RS, Visscher PK, Camazine S., Mass envenomations by honey bees and wasps, in West J Med 1999 Apr;170(4):223-7
Vlasak R, Kreil G., Nucleotide sequence of cloned cDNAs coding for preprosecapin, a major product of queen-bee venom glands, in Eur J Biochem 1984 Dec 3;145(2):279-82
 |
||||||||||||
| LU | LU BF | LDF | Ezeri | Sugu enciklopÄdija |
BioloÄĢiskÄ daudzveidÄŦba |
Latvijas daba | Piekrastes biotopi |
Malokologu biedrÄŦba |
BotÄnikas biedrÄŦba |
MeklÄ | Raksti mums |