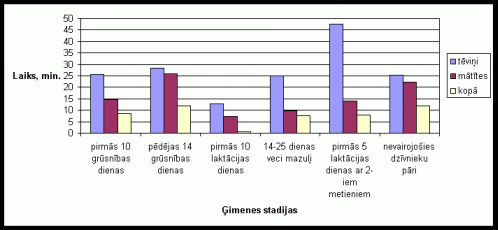
1. attįls Vidįjais ârpus migas pavadîtais laiks vienâ aktivitâtes ciklâ Íînas strupastes tįviōiem un mâtîtįm atseviðíi un kopâ, atkarîbâ no ėimenes stadijas.
(uz tekstu)
Teksts: Oskars Rozenkopfs
Noformįjums: Edgars Erâns
Biol B 97045
sb70045@lanet.lv
ÍÎNAS STRUPASTU Lasiopodomys mandarinus, KÂ MONOGÂMAS SUGAS, UZVEDÎBAS ÎPATNÎBAS
Saturs
Ievads
1. Literatûras
apskats
1.1. Íînas
strupastu raksturojums
1.1.1. Sistemâtika
1.1.2. Izplatîba
1.1.3. Dzîvesveids
1.1.4. Vairoðanâs
1.1.5. Ekoloėija
1.2. Zîdîtâju
sociâlo sistįmu un uzvedîbas veidi
1.2.1. Zîdîtâju
sociâlo sistįmu veidi
1.2.2. Monogâmijas
sastopamîba zîdîtâjiem
1.2.3. Monogâmiju
raksturojoðie faktori
1.2.4. Vecâku
uzvedîbu raksturojoðie sociâlie faktori
1.2.5. Hormonâlâ
ietekme uz vecâku uzvedîbu
2. Materiâls
un metodika
Rezultâti
Diskusijas
Secinâjumi
Literatûras
saraksts
Neskatoties uz to, ka zîdîtâju uzvedîba ir diezgan plaði pįtîta, ðajâ jomâ pastâv diezgan daudzas neskaidrîbas. Îpaði tas sakâms par monogâmo dzîvnieku uzvedîbas izpįti.
Par monogâmijas galveno râdîtâju tiek uzskatîts sociâlo pâra saiðu veidoðanâs starp viena pâra tįviōu un mâtîti, kas ilgst vismaz lîdz viena mazuïu metiena izaudzinâðanai un parasti vįl ilgâk (Dewsbery 1981). Monogâmiem zîdîtâjiem ir raksturîga ėimenes veidoðana. Ėimene var bût parasta, viens vecâks vai pârojoðais pâris, kas audzina mazuïus, kâ arî paplaðinâtâ ėimene, kurâvar bût viens vai vairâki tâ paða dzimuma radinieki. Tįviōa klâtbûtne nav nepiecieðama, lai kâda sociâlâ grupa tiktu uzskatîta par ėimeni (Emlen 1995). Dzîvnieku izpįte ðâdâ veidâ paver mums vįrtîgu logu, caur kuru mįs varam vieglâk vįrot fundamentâlâs bioloėiskâs likumsakarîbas, kas vada sociâlâs mijiedarbîbas ėimeōu grupâs. Veroties ðajâ logâ mums tiek dots ieskats ne-kultûras faktoros, kas nosaka mûsu paðu uzvedîbu (Emlen 1995). Veicot ðâdus pįtîjumus ir ïoti interesanti aplûkot seksuâlâs un sociâlâs monogâmijas atðíirîbas. Monogâmie dzîvnieki, tâpat kâ cilvįki ne vienmįr stâjas seksuâlajâs attiecîbâs tikai ar vienu partneri. Gan monogâmie dzîvnieki, gan cilvįki ir daudz izvįlîgâki izvįloties sociâlo nevis seksuâlo partneri. Pįtîjumi par ðâdâm monogâmajâm ėimeōu grupâm ir veikti salîdzinoði maz, tâpįc ir nepiecieðams uzzinât par ðâdu monogâmo dzîvnieku uzvedîbu un pieauguðo îpatōu lomu ėimenes dzîvį. Par izpįtes objektu tika izmantoti Lasiopodomys mandarinus sugas pârstâvji, kuriem ir raksturîga gan monogâmijas gan ėimenes saiðu. Monogâmija ir reti sastopama zîdîtâjiem un monogâmâs attiecîbas seksuâlâs uzvedîbas sfįrâ nav pietiekami pįtîtas arî Íînas strupastįm (Zakharov et al. 1998).
Pįtîjums mįríis ir izpįtît ðîs sugas tįviōu un mâtîðu ieguldîjumu ėimenes funkciju uzturįðana daþâdâs stadijâs. Ðim darba mįríim ir izveidoti sekojoði darba uzdevumi: noskaidrot tieðo un netieðo vecâku rûpju intensitâti mâtîtįm un tįviōiem; noskaidrot mâtîðu un tįviōu dalîbu teritorijas aizsardzîbâ; noskaidrot tįviōa partnera aizsardzîbas no pâroðanâs ar sveðiem tįviōiem, esamîbu.
Íînas strupastes apdzîvo tipčaka stepes, ðíembainâs un akmeōainâs vietâs starp stepju pïaviōâm . Selengijas un Djuidinskovas rajonâ strupastes izvairâs no augstzâïu rajoniem un ir satopami tikai tipčaka stepįs, kur ir auga Stellera chamaejosme pârpilnîba. Uz R no Armaka rajona L. mandarinus alas ir atrastas gan tipčaka stepįs, gan stepįs ar bagâtîgu pïavu graudzâïu veėetâciju, retos gadîjumos ðî suga sastopama arî tîrumos ( 1997).
1.2.Zîdîtâju sociâlo sistįmu un uzvedîbas veidi
1.2.1.Zîdîtâju sociâlo sistįmu veidi
Atseviðíâm zîdîtâju sugâm ir raksturîga ėimeōu veidoðana. Ėimenes ir kopâ dzîvojoðas îpatōu grupas, kuras ir spįjîgas radît, apmâcît un sociâli izaudzinât mazuïus (Good 1964; Ferber 1964; Nock 1992; cit. pįc Emlen 1995). Ėimene sastâvâ ietilpst vismaz vecâki un seksuâlo dzîvi neuzsâkuðie îpatōi (Fox 1967; van den Berghe 1979; cit. pįc Emlen 1995). Ėimenįm var bût raksturîga nepatstâvîba, tâs var tikt veidotas un attîstîtas, kad ir nepiecieðamas (lîdz mazuïu nobrieðanai) (Emlen 1995). Ðâdâm ėimenįm ir raksturîgas kolektîvas rûpes par mazuïiem (Emlen 1991, cit. pįc Konig 1994). No ~4400 zîdîtâju sugâm, kas tika pârbaudîtas, ėimenes veidoðanas iezîmes izrâdîja tikai 15% no tâm. Sociâlâ sistįma, kurâ mazuïus zîda dominįjoða mâtîte un abu dzimumu dzîvnieki rûpįjas par viōas mazuïiem, ir raksturîga, piemįram, ðâdâm sugâm Lycaon pictus, Canis aureus, Canis mesomelas, Helogale parvula, Suricata suricatta, Hetreroceptalus glaber. Ir raksturîgas arî grupas, kurâs mâtîtes kopîgi zîda un kopj mazuïus, piemįram Panthera leo, Crocuta crocuta, Nasua nasua, Mungos mungo. (Konig 1994). Bara locekïu rûpes par mazuïiem, kas nav viōu pįcnâcįji, varįtu uzskatît par altruistiskâm (Hamilton 1963, cit. pįc Konig 1994). Tomįr ðâdas rûpes sniedz tiem sociâlo pieredzi un var noderįt rûpįjoties par saviem mazuïiem (Konig 1994)
1.2.2.Monogâmijas sastopamîba zîdîtâjiemMonogâmija, kas ir raksturîga putniem, ir reti sastopama zîdîtâjiem (Carter, Getz Lowell 1993). Pįc Kleimanes (Kleiman) datiem tikai aptuveni 3% zîdîtâjiem piemît monogâms dzîvesveids (cit. pįc Carter, Getz Lowell 1993). Monogâmas ir atseviðías plįsįju, primâtu, grauzįju u.c. kârtu sugas. Monogâmiju izrâda tâdas atðíirîgas sugas, kâ savvaïas suōi, tamarîni un murkðíi (Carter,Getz Lowell 1993). Djûsberijs (Dewsbury), veicot eksperimentus ar grauzįjiem, izveidoja tâ saucamo monogâmijas skalu. No eksperimentâ 30 iesaistîtajâm sugâm tikai septiōas ðajâ skalâ ieguva pozitîvu râdîtâju; Peromyiscus californicus, Onychomys torridus, Peromyscus polionotus, Peromyscus eremicus, Peromyscus polionotus, Onychomys leucogaster, Ochrotomys nuttalli (Dewsbury 1981). Pie sugâm, kas tika iekïautas ðajâ eksperimentâ, pie monogâmiem netika iekïautas Microtus pinetorum un Microtus ochrogaster, kas arî tiek pieskaitîtas pie monogâmajiem grauzįjiem (Carter, Getz Lowell 1993). Arî L. mandarinus ir monogâma grauzįju suga. (Zakharov et al.1998).
1.2.3.Monogâmiju raksturojoðie faktoriMûsdienu uzvedîbas ekologi uzskata, ka monogâmija ir radusies tâpįc, ka atseviðíâm sugâm raduðies poligînijai nelabvįlîgi apstâkïi (Krebs, Davies 1993, cit. pįc Reyer 1994). Lîdz ar to rodas it kâ pretrunas starp abu dzîvnieku interesįm. Tįviōi cenðas savas mâtîtes nosargât no pâroðanâs ar citiem tįviōiem, bet mâtîtes cenðas sapâroties ar citiem tįviōiem (Reyer 1994). Galvenais kritįrijs, kas raksturo monogâmiju ir ilgstoðas attiecîbas starp tįviōu un mâtîti (Carter, Getz Lowell 1993). Par monogâmiem tiek uzskatîti dzîvnieki, kuriem saglabâjas pâra saites vismaz no pâroðanâs sâkuma lîdz mazuïu izaudzinâðanas beigâm (Dewsbury 1981). Monogâmijas sociâlai organizâcijai parasti raksturîga rûpîga partnera izvįle, spįcîgi attîstîtas partnera jûtas un augsta agresivitâte pret sveðiniekiem, aizsargâjot teritoriju, migu un partneri (Carter, Getz Lowell 1993; Insel et al. 1995). Monogâmo sugu tįviōi un mâtîtes parasti ir lîdzîgi pįc izmįra un izskata (Dewsbury 1981; Carter, Getz Lowell 1993). Monogâmo sugu dzîvniekiem ir raksturîgs garð latentais periods (Zakharov et al. 1998). Monogâmajiem dzîvniekiem ir raksturîga bieþâka sociâlâ kontaktįðanâs un rûpes vienam par otru nekâ nemonogâmajiem dzîvniekiem (Dewsbury 1981; Carter, Getz Lowell 1993). Monogâmo zîdîtâju tįviōiem ir raksturîgs mazs ejakulâciju skaits viena mâtîtes estrusa laikâ salîdzinot ar nemonogâmo sugu tįviōiem (Dewsbury 1981; Carter, Getz Lowell 1993). Monogâmo sugu tįviōi veic atkârtotu pâroðanos ar mâtîti, ja tiem ir aizdomas par mâtîtes pâroðanos ar citu tįviōu (Dewsbury 1981; Zakharov et al. 1998). Ðim pielâgojumam varįtu bût tįviōa ėenįtisko materiâlu aizsargâjoða funkcija (Dewsbury 1981). Daudzu sugu tįviōiem pârojoties ar vienu mâtîti rodas seksuâlais pârsâtinâjums. Ðie tįviōi, bieþi vien, normâlu vairoðanâs spįju var atgût vairojoties ar citu mâtîti. Ðo parâdîbu sauc par Kûlaidþa efektu (Coolidge effect), taču to sugu tįviōiem, kuriem raksturîga monogâmija, partneru maiōa izrâdîja mazâku efektivitâti nekâ sugâm, kurâm raksturîgi poligînija vai gadîjuma dzimumsakari (Dewsbury 1981). Monogâmo sugu grauzįjiem ir raksturîgs arî zems reproduktîvais potenciâls salîdzinot ar citâm grauzįju sugâm (Dewsbury 1981). Piemįram, Microtus pinetorum (monogâma suga) 2.3 mazuïi metienâ, bet Rattus rattus (poligâma suga) 6.6 mazuïi metienâ (Dewsbury 1981). Monogâmo sugu dzîvnieku mazuïiem ir raksturîga salîdzinoði lįna individuâlâ attîstîba (Dewsbury 1981). Piemįram, acu atvįrðanâs mazuïiem Microtus pennsylvanicus (poligâma suga) notiek 9.4 dienâs, bet Onychomys torridus (monogâma suga) 15 dienâs. Vįl viena raksturîga monogâmo dzîvnieku pazîme ir vįla seksuâla nobrieðana (Dewsbury 1981). Pârsvarâ gadîjumu monogâmo dzîvnieku pįcnâcįji nesasniedz pubertâti tik ilgi, kamįr dzîvo kopâ ar vecâkiem (Carter, Getz Lowell 1993). Monogâmiem dzîvniekiem nav novįrojama pâroðanâs tuvu asinsradinieku starpâ (Carter, Getz Lowell 1993). Veicot novįrojumus daþâdu monogâmu sugu strupastįm, tika novįrots, ka, lai arî to dzîvesveids ir monogâms, tomįr ðie dzîvnieki veido seksuâlâs attiecîbas ne tikai ar savu partneri (Dewsbury 1981; Zakharov et al. 1998). Veicot DNS testus sugas Microtus ochrogaster mazuïiem, tika konstatįts, ka mâtîðu pįcnâcįju tįvi ne vienmįr ir tie tįviōi ar kuriem tâs dzîvo kopâ, bieþi vien mazuïiem tika konstatįta jaukta paternitâte. Lîdz ar to var redzįt, ka sociâlâ monogâmija ne vienmįr nozîmį pilnîgu seksuâlo monogâmiju (Carter, Getz Lowell 1993). Pie sociâlâs monogâmijas ïoti svarîga ir pâru saiðu veidoðanâs. Ðîs pâru saites galvenokârt veidojas pârojoties. Îpaði svarîga M. ochrogaster mâtîtįm ir pirmâ pâroðanâs reize, kas ir ïoti ilgstoða un kuras laikâ veidojas sociâlâs pâra saites uz visu mûþu. Mâtîte pirmâs pâroðanâs reizį var bût uzbudinâta 30-40 stundas un ðajâ laikâ notiek atkârtota vairâkkârtįja pâroðanâs (Carter, Getz Lowell 1993).
1.2.4.Vecâku uzvedîbu raksturojoðie sociâlie faktoriPįtot monogâmos zîdîtâjus tika atklâti daudzi sociâlie faktori, kas raksturîgi vecâku uzvedîbai. Ïoti daudzu monogâmo sugu zîdîtâji pastiprinâtu agresiju pret sveðiem dzîvniekiem un savas teritorijas aizsargâðanu sâk izrâdît jau pįc pâroðanâs (Carter, Getz Lowell 1993; Insel 1994 u.c.). Ðî agresivitâte un teritorijas aizsargâðana ir ïoti izteikta pįc mazuïu piedzimðanas un laktâcijas periodâ (Carter, Getz Lowell 1993). Monogâmo sugu zîdîtâju uzvedîbai ir raksturîgas ðâdas sociâlâs iezîmes: mazuïu sildîðana, baroðana (mâtîtįm), apkopðana, pârnįsâðana, kontaktįðanâs ar mazuïiem. Ðîs rûpes ir raksturîgas abiem vecâkiem, taču galvenokârt mâtîtįm. Tįviōi ïoti daudz laika pavada ârpus migas, sargâjot un iezîmįjot teritoriju, taisot migu un rokot (McGuire, Novak 1984; Solomon, 1993). Monogâmo sugu tįviōiem ir raksturîgi atraðanâs migâ brîdî, kas mâtîte atrodas ârpus migas (McGuire, Solomon 1984). Ïoti liela nozîme ir arî vecâku sociâlajai pieredzei. Veicot eksperimentus ar M. ochrogaster tika konstatįts, ka pieredzįjuðas mâtîtes pavadîja mazâk laika ârpus miga, bet vairâk laika, aprûpįjot mazuïus. Ðo mâtîðu mazuïi attîstîjâs âtrâk un to izdzîvoðanas koeficients bija daudz augstâks, un svars vecumâ, kad tie kïuva patstâvîgi, bija lielâks nekâ nepieredzįjuðu mâtîðu mazuïiem. Mazuïu kopðanas pieredze neizrâdîja nekâdu ietekmi uz tįviōu uzvedîbu (Wang, Novak 1994). Pįtot monogâmo peïu sugu Peromyscus californicus tika konstatįts, ka tįviōiem, kuriem nav iepriekðįja vecâku pieredze, ir nepiecieðama mâtîtes klâtbûtne, lai tie izrâdîtu vecâku rûpes un nenogalinâtu mazuïus. Savukârt, tįviōi ar iepriekðįju vecâku pieredzi izrâdîja rûpes par mazuïiem arî bez mâtîtes klâtbûtnes (Gubernick 1993).
1.2.5.Hormonâlâ ietekme uz vecâku uzvedîbuVeicot eksperimentu ar M. ochrogaster pįtnieki atklâja, ka ïoti nozîmîga uz monogâmijas un vecâku rûpju attîstîbu ir hormonâlâ ietekme. Ïoti iespįjams, ka viens no galvenajiem sociâlo saiðu nostiprinoðajiem faktoriem ir augstais glikokortikoîdu saturs monogâmo dzîvnieku asinîs, ko izraisa pastiprinâta virsnieru dziedzeru aktivitâte. Virsnieru dziedzera aktivitâte nosaka arî samazinâtu seksuâlo dimorfismu un palielinâtas vecâku rûpes tįviōiem. Tika atklâts, ka jau neilgi pįc piedzimðanas M. ochrogaster mazuïiem ir neparasti aktîva virsnieru dziedzeru sistįma. Izolįjot mâti uz daþâm minûtįm bįrniem spįcîgi paaugstinâs glikokortikoîdu lîmenis asinîs. Eksperimenti ir pierâdîjuði, ka pastiprinâta virsnieru dziedzeru aktivitâte arî poligâmo sugu dzîvniekiem izraisa lîdzîgu vecâku uzvedîbu, kâ monogâmo sugu dzîvniekiem (Carter, Getz Lowell 1993). Milzîga nozîme monogâmijas, vecâku rûpju veidoðanâ un pâra saiðu nostiprinâðanâ ir hormoniem oksitocînam un vazopresînam. Abu ðo hormonu izdalîðanâs notiek pie lîdzîgiem apstâkïiem seksuâlo un citu kontaktu starpâ. Tika novįrots, ka oksitocîna pastiprinâta izdalîðanâs notiek, ja tiek kairinâti mâtîtes zîdekïi kâ arî ėenitâlijas. Arî vazopresîns strauji izdalâs pâroðanâs laikâ un pįc tâs. Ðo abu hormonu funkcijas ir antagoniskas, ja oksitocîns izraisa sociâlu uzvedîbu, tad vazopresîns izraisa antisociâlu uzvedîbu. Dzîvniekiem, kas dzîvo kopâ, oksitocîna pastiprinâta izdalîðanâs ir novįrota, pat pie vienkârðas saskarðanâs. Injicįjot mâtîtįm oksitocînu CNS, tâs kïûst sociâlâkas un mazâk agresîvas salîdzinot ar mâtîtâm, kuras nav saōįmuðas oksitocîna injekciju, vai mâtîtįm, kuras saōįmuðas ðo injekciju perifįrija asinsritį. Mâtîtįm, kurâm ir injicįts oksitocîns, âtrâk veidojas seksuâlâs attiecîbas ar tįviōiem.( Carter, Getz Lowell 1993). M. ochrogaster mâtîtes bez seksuâlâs pieredzes sâk pâroties tikai divas nedįïas pįc kopdzîves ar tįviōu. Ðîm mâtîtįm injicįjot oksitocînu pâroðanâs tika novįrota daudz âtrâk. Ja pįc tam mâtîtįm injicįja oksitocîna antagonistus, tâm atkal parâdîjâs normâla seksuâlâ uzvedîba. Injicįjot okstitocîna antagonistus mâtîtįm ar seksuâlu pieredzi, ðî injekcija nespįja ietekmįt mâtîtes sociâlo uzvedîbu pret tįviōu (Insel, Thomas 1995). Pįc Insela (Insel) novįrojumiem oksitocîns ir galvenais sociâlâs uzvedîbas noteicįjs. Injicįjot oksitocîna antagonistus CNS tika konstatįs, ka mâtîtįm ir traucįtas visas sociâlâs funkcijas, izōemot pâroðanos. Vįl vairâk, injicįjot oksitocînu, M. ochrogaster veidoja sociâlâs saites pat bez pâroðanâs (Insel et al., 1995). Ïoti svarîgs ir arî oksitocîna receptoru izvietojums CNS. Monogâmo sugu dzîvniekiem tas atðíiras no nemonogâmo sugu dzîvniekiem.( Carter, Getz Lowell 1993; Insel et al. 1995; Wang, Novak 1996). Monogâmiem dzîvniekiem ir augsti attîstîta vecâku uzvedîba, kâ arî izteikta savas ėimenes, partnera un teritorijas aizsargâðana un agresija pret sveðiem dzîvniekiem. Pįtnieki ir secinâjuði, ka ðo izpausmju galvenais izraisîtâjs ir vazopresîns (Carter, Getz Lowell 1993).
Pįtîjumos tika izmantota suga Íînas strupaste (Lasiopodomys mandarinus). Pįtîjumâ iesaistîtie dzîvnieki ir dzimuði Rîgâ vairâkâs paaudzįs, savukârt to senči tika ievesti no Sanktpįterburgas, uz kurieni tos atveda no Burjatijas republikas Selengijas rajona, kur tie tika noíerti to dabîgajâs dzîves vietâs.
Lîdz pįtîjumiem dzîvnieki tika turįti 43*28*15 cm lielos plastmasa bûros ar stiepïu pârsegumu 19 +/- 3 °C temperatûrâ. Laboratorijas apgaismojums ir 18h diennaktî, ðâds fotoperiods ir ïoti labvįlîgs priekð grauzįju vairoðanâs (Ferkin, Kile 1996; Baker, Clarke 1989). Laboratorijâ ir neliels logs, pa kuru iespîd gaisma, kas var izmainît fotoperiodisma sadalîjumu. Ðobrîd laboratorijas dzîvniekiem barîbâ dod sauso barîbu kâ arî ûdeni (neierobeþotâ daudzumâ) (Baker, Clarke 1989). Tâ kâ materiâlais stâvoklis to neatïauj un Íînas strupastįm nav izstrâdâtas speciâlas baroðanas devas, tad par barîbu tika izmantoti: burkâni (neierobeþotâ daudzumâ), âboli, auzu pârslas Herkuless, mâtîðu grûtniecîbas un laktâcijas periodâ nedaudz saulespuíu sįklu, vasarâ un pavasarî tika dotas nedaudz pieneōu lapu, bet, sâkot ar rudens beigâm lîdz pavasarim (pirms pieneōu lapu parâdîðanâs), tika dots nedaudz auzu dîgstu. Kâ pakaiði tika izmantotas koka įveïskaidas (apmįram trîs centimetru biezumâ), bet kâ migas materiâls- siens.
Reėistrįjot un analizįjot rezultâtus, tika ōemti vįrâ ðâdi uzvedîbas veidi: ârpus migas pavadîtas laiks tįviōam un mâtîtei atseviðíi, kâ arî kopâ ârpus migas pavadîtai laiks, vidįjais un procentuâlais rakðanas laiks ârpus migas, vidįjais migas materiâla neðanas skaits vienâ stundâ, vidįjais uzrâpðanâs reiþu skaits uz íieėeïiem vienâ stundâ kâ arî uz íieėeïiem pavadîtais laiks, vidįjais urinįðanas reiþu skaits uz íieėeïiem vienâ stundâ, pârskrįjienu un teritorijas apgaitas skrįjienu skaits vienâ stundâ, uzmanîbas pozu skaits vienâ stundâ un tajâs pavadîtais laiks, nazo-nazâlo un nazo-anâlo kontaktu skaits vienâ stundâ, vidįjais un procentuâlais įðanas laiks ârpus migas. Par pârskrįjieniem tika uzskatîti strauji, spįji, aptuveni 30 cm gari skrįjieni, kuru laikâ dzîvnieku aste bija pacelta. Par teritorijas apgaitas skrįjieniem tika uzskatîti apïveida skrįjieni, kas aptvįra visu teritoriju. Arî ðo skrįjienu laikâ dzîvnieku aste bija pacelta. Par uzmanîbas pozu tika uzskatîta dzîvnieku poza, kad tie bez kâda ârįja iemesla sastinga uz vietas ar izvįrstâm priekðíepâm, pieglaustâm ausîm un paceltu asti.
Ðie aktivitâtes veidi tika izvįlįti tâpįc, ka priekðnovįrojumos tika konstatįta to sastopamîba gandrîz visos ėimenes stâvokïos, turklât to aktivitâte daþâdos ėimenes stâvokïos atðíîrâs. Atseviðíi no ðiem uzvedîbas veidiem arî raksturo tįviōa un mâtîtes tieðâs un netieðâs vecâku rûpes par mazuïiem. Piemįram, migas materiâla un barîbas neðana uz migu ir pieskaitâmas netieðajâm vecâku rûpįm, bet mazuïu laizîðana, neðana un vâkðana- tieðajâm vecâku rûpįm (Xia, Millar 1987). Tâ kâ novįrojumi migas iekðienį netika veikti, tad par tieðajâm vecâku rûpįm tika uzskatîts viss laiks, ko dzîvnieki pavadîja migâ pie mazuïiem. Pie netieðajâmvecâku rûpįm tika pieskaitîta arî pakaiðu rakðana, kas aizvieto dabâ sastopamo alu un tuneïu veidoðanu. Par migas aizsardzîbas un partnera aizsardzîbas no pâroðanâs aktivitâtįm tika uzskatîta uzrâpðanâs uz íieėeïiem un urinįðana uz tiem pârskrįjieni un teritorijas apgaitas skrįjieni, uzmanîbas pozas kâ arî nazo-anâlie kontakti. Nazo-nazâlie kontakti un daïa nazo-anâlo kontaktu tika vįrtįta kâ atpazîðanas uzvedîba. Ârpus migas pavadîtais laiks un įðanai ârpus migas veltîtais laiks tika izmantots vispârįjâs dzîvnieku ârpusmigas aktivitâtes raksturoðanai, attiecîgajâ ėimenes stadijâ. Ârpus migas aktivitâte tika uzskaitîta, ja novįrotais îpatnis pirms aktivitâtes cikla migâ bija pavadîjis vismaz desmit minûtes. Tâpat arî, ja viena novįrojama laikâ tįviōð vai mâtîte starp aktivitâtes periodiem migâ pavadîja vismaz desmit minûtes, katrs aktivitâtes periods tika uzskatîts par atseviðíu aktivitâtes ciklu. Par katra îpatōa atseviðías aktivitâtes ciklu tika uzskatîts aktivitâtes periods, kad mâtîte vai tįviōð pavadîjaârpus migas vismaz trîs minûtes, jo priekðnovįrojumos tika konstatįts, ka dzîvnieki mįdz iznâkt ârpus migas arî pasivitâtes cikla laikâ uz samįrâ neilgu laika brîdi, kura laikâ neizrâda aktivitâtes ciklam raksturîgu uzvedîbu. Par tįviōa un mâtîtes kopįjo aktivitâtes ciklu tika uzskatîts kopįjâs aktivitâtes periods, kura laikâ mâtîte un tįviōð kopîgi pavadîja ârpus migas vismaz 30 sekundes. Ðâds laiks par kopįju aktivitâtes cikla laiku tika pieōemts tâpįc, ka dzîvnieku atraðanâs ârpus migas tika novįrota arî îsâkos laika periodos, taču bieþi vien, îpaði laktâcijas laikâ, pįc otra dzîvnieka parâdîðanâs ârpus migas, viens no tiem atgriezâs migâ un dzîvnieki neizrâdîja kopîgu aktivitâti.
Ōemot vįrâ literatûrâ atrodamos datus un novįrojumu laikâ iegûtâs atziōas par Íînas strupastu uzvedîbu, ar ðiem dzîvniekiem tika veikti arî vairâki eksperimenti.
Tâ kâ Íînas strupaste ir pieskaitâma pie monogâmâm sugâm, tad varįtu paredzįt teritorijas aizsardzîbas uzvedîbas pastiprinâðanos ėimenes pâru locekïiem, konstatįjot savâ teritorijâ sveðu tįviōu vai tâ klâtbûtnes pįdas, tâpat arî tįviōa uzvedîbas pastiprinâðanos, kas vįrsta uz mâtîtes aizsardzîbu no pâroðanâs ar sveðu tįviōu tâs seksuâlâs aktivitâtes laikâ. Ðî paredzįjuma pârbaudîðanai ilgstoði kopâ dzîvojoðiem tįviōiem un mâtîtįm tika dota sveða seksuâli aktîva tįviōa smarþa. Ðajâ nolûkâ ðî ėimenes pâra teritorijâ tika ieliktas skaidas, no ðâda tįviōa bûra, samitrinâtas ar tâ urînu. Skaidas bûrî tika ievietotas dzîvniekiem nemanot. Uzvedîbas reėistrâcija tika sâkta no tâ brîþa, kad kâds no ėimenes pâra dzîvniekiem ievįroja skaidas un tâs apoðōâja un tika reėistrįta lîdz brîdim, kad vismaz viens no pâra locekïiem iegâja atpakaï migâ. Eksperimentâ tika izdalîtas divas dzîvnieku grupas. Pirmajâ grupâ tika iekïauti seksuâli nobrieduðu dzîvnieku pâri, kuros mâtîtes visticamâk atradâs diestrusa stâvoklî (novįrojuma dienâ un 2 lîdz 3 dienas pirms novįrojuma netika konstatįti nekâdas seksuâlas aktivitâtes uzvedîba). Otru grupu sastâdîja dzîvnieku pâri, kuros mâtîtes atradâs seksuâlas aktivitâtes stâvoklî (sprieþot pįc dzîvnieku uzvedîbas bûrî) vai vienu lîdz divas dienas lîdz estrusam (grûsnas mâtîtes vienu lîdz divas dienas lîdz dzemdîbâm). Pirmâs grupas dzîvnieki sastâdîja piecus pârus un ar tiem tika izdarîti astoōi novįrojumi, otrâs grupas dzîvnieki sastâdîja astoōus pârus un tika izdarîti deviōi eksperimenti. Kopîgais novįrojumiem veltîtais laiks bija apmįram septiōas stundas.
Íînas strupastįm ir raksturîgas augstas vecâku rûpes kâ arî rûpes par teritoriju un fona novįrojumu laikâ tâm tika konstatįta samįrâ augsta migas materiâla neðanas aktivitâte, tad tika nolemts veikt eksperimentus konkrįti ðim uzvedîbas veidam. Dzîvniekiem neredzot, bûrî tika ievietots papildus migas materiâls un novįrojumu reėistrâcija tika sâkta tad, kad kâds no pâra dzîvniekiem sâka nest ðo migas materiâlu uz migu. Novįrojums tika turpinâts tik ilgi, kamįr vismaz viens no pâra dzîvniekiem iegâja atpakaï migâ. Novįrotie dzîvnieku pâri tika sadalîti divâs grupâs dzîvnieku pâri ar mazuïiem un dzîvnieku pâri bez mazuïiem. Tâ kâ eksperimentu skaits bija neliels, tad otrajâ dzîvnieku grupâ tika iekïauti arî pâri, kuru mâtîtes bija grûsnas, lîdz ar to ðo pâru uzvedîba varįja atðíirties no nepârojoðos dzîvnieku pâru uzvedîbas. Pirmajâ dzîvnieku grupâ tika iekïauti divi dzîvnieku pâri un ar tiem tika izdarîti četri novįrojumi. Otrajâ dzîvnieku grupâ iekïâva piecus dzîvnieku pârus un ar tiem tika izdarîti septiōi novįrojumi. Kopumâ novįrojumi ar ðiem dzîvniekiem tika veikti ~13 stundas.
Novįrojumi tika izdarîti pįc iespįjas netraucįjot dzîvniekus. Dati tika pierakstîti kladį ar speciâli ieviestu zîmju palîdzîbu. Uzvedîbas aktivitâðu fiksįðanai, kuru raksturoðanai bija nepiecieðams laiks, tika izmantots hronometrs.
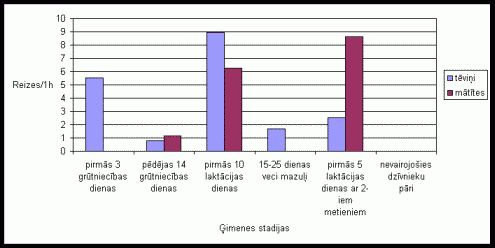
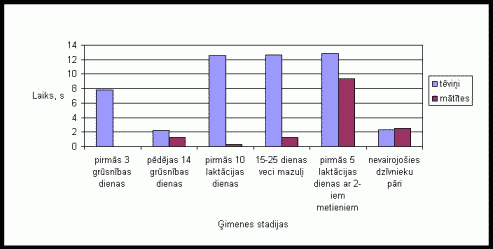
Apskatot tįviōu un mâtîtes ârpus migas pavadîto laiku var novįrot ðâdus rezultâtus. Lai arî visumâ abu îpatōu aktivitâte bija lîdzîga, tomįr novįrojams, ka ârpus migas tįviōð pavadîja lielâku laiku nekâ mâtîte (skat. 1. att. ). Îpaði augstu aktivitâti tįviōð uzrâdîja pirmajâs 10 grûtniecîbas dienâs dzîvojot kopâ ar 12-25 dienas veciem mazuïiem un dzîvojot kopâ ar divu metienu mazuïiem, kâ arî pįdįjâs 14 grûtniecîbas dienâs. Mâtîte visaugstâko aktivitâti izrâdîja pįdįjâs 14 grûtniecîbas dienâs un dzîvojot kopâ ar nevairojoðiem dzîvnieku pâriem. Kopîgâ aktivitâte dzîvniekiem visaugstâkâ bija pįdįjâs 14 grûtniecîbas dienâs un dzîvojot kopâ ar nevairojoðiem dzîvnieku pâriem. Pirmajâs 10 laktâcijas dienâs tįviōu un mâtîðu kopîgâ aktivitâte gandrîz netika novįrota.
1. attįls Vidįjais
ârpus migas pavadîtais laiks vienâ aktivitâtes ciklâ Íînas strupastes tįviōiem
un mâtîtįm atseviðíi un kopâ, atkarîbâ no ėimenes stadijas.
(uz
tekstu)
Rakðanâs skaidâs, kas varįtu bût ekvivalenta dabâ sastopamai alu rakðanai un ir pieskaitâma netieðajâm rûpįm par mazuïiem, bija raksturîga abiem îpatōiem. Visumâ bija vįrojams, ka gan procentuâli, gan vidįji tįviōð rokot pavada vairâk laika (skat. 2. att.). Vislielâkâ rakðanas aktivitâte tįviōam bija vįrojama dzîvojot kopâ ar 12-25 dienas veciem mazuïiem. Samįrâ augsta aktivitâte bija vįrojama arî pirmajâs 10 laktâcijas dienâs, pįdįjâs 14 grûtniecîbas dienâs un dzîvojot kopâ ar divu metienu mazuïiem. Mâtîtei visaugstâkâ rakðanas aktivitâte bija vįrojamâ dzîvojot kopâ nevairojoðos dzîvnieku pâriem, kâ arî pirmajâs 10 laktâcijas dienâs.
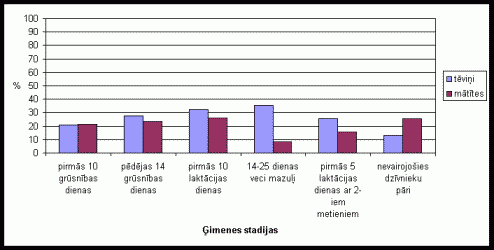
2. attįls Rakðanas
procentuâlais laiks (no kopįjâ aktivitâtes laika) Íînas strupastes tįviōiem
un mâtîtįm atseviðíi, atkarîbâ no ėimenes stadijas.
(uz tekstu)
Migas materiâlu neðanas aktivitâte bija samįrâ lîdzîga visu ėimenes stadiju laikâ (skat. 3. att.). Migas materiâlu neðanas pieaugums mâtîtei bija raksturîgs dzîvojot kopâ ar divu metienu mazuïiem. Ïoti zema migas materiâlu neðanas aktivitâte bija vįrojama dzîvojot kopâ nevairojoðiem dzîvnieku pâriem. Eksperimentos, kad tika pievienots papildus migas materiâls, bija novįrojama augsta migas materiâlu neðanas aktivitâte (skat. 4. att.). Ïoti augstu aktivitâti izrâdîja tįviōi. Îpaði augsta aktivitâte tįviōiem bija novįrojama dzîvojot kopâ ar mazuïiem.
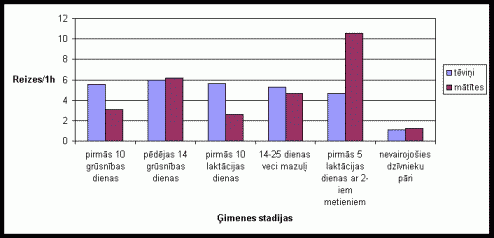
3. attįls Migas materiâla
neðanas skaits vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem un mâtîtįm
atkarîbâ no ėimenes stadijas.
(uz tekstu)
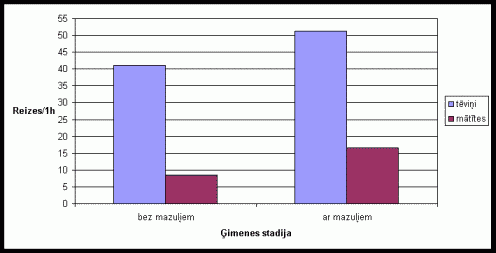
4. attįls Migas materiâla
neðanas skaits vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem un mâtîtįm
atkarîbâ no ėimenes stadijas, ja pirms eksperimenta bûrî ievietots svaigs
migas materiâls.
(uz tekstu)
Aplûkojot datus, kas iegûti reėistrįjot dzîvnieku urinįðanu uz íieėeïiem (skat. 5. att.), varįja novįrot, ka tįviōiem visaugstâkâ urinįðanas aktivitâte bija dzîvojot kopâ ar 12-25 dienas veciem mazuïiem un dzîvojot kopâ ar nevairojoðos dzîvnieku pâriem. Mâtîtįm visaugstâkâ urinįðanas aktivitâte tika novįrota dzîvojot kopâ ar 12-25 dienas veciem mazuïiem, savukârt ïoti zema visasgrûtniecîbas laikâ. Mâtîtįm bija raksturîga urinįðana lielu lâðu veidâ, savukârt tįviōiem nelielu urîna strîpiōu (iegarenu pilienu) vai pilienu virkōu veidâ. Eksperimentos, kad dzîvnieku bûrî tika ievietotas skaidas ar sveða tįviōa smarþu, bija novįrojama strauja urinįðanas aktivitâtes palielinâðanâs (skat. 6. att.). Dzîvnieku pâriem, kuros mâtîtes atradâs diestrusa stâvoklî (1. grupa), augstâku urinįðanas aktivitâti uzrâdîja mâtîtes, savukârt pâros, kuros mâtîtes atradâs estrusa vai pirmsestrusa stâvoklî (2. grupa), augstâku urinįðanas aktivitâti uzrâdîja tįviōð.
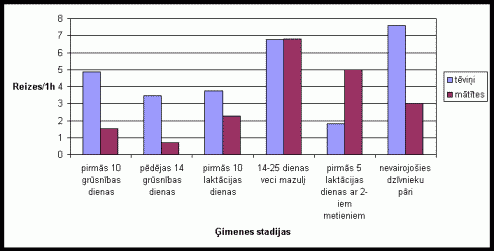
5. attįls Urinįðanas
skaits uz íieėeïiem vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem un
mâtîtįm atkarîbâ no ėimenes stadijas.
(uz tekstu)
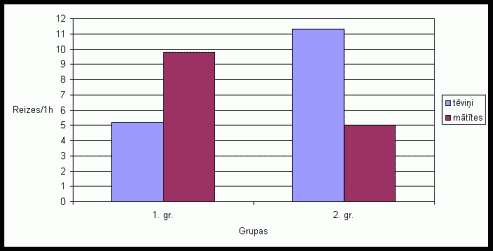
6. attįls Urinįðanas
skaits uz íieėeïiem vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem un
mâtîtįm, ja pirms eksperimenta bûrî tika ievietotas skaidas ar sveða tįviōa
smarþu.
(uz tekstu)
Augstu aktivitâti dzîvnieki uzrâda, uzrâpjoties uz íieėeïiem (skat. 7. att.). Aptuveni vienâdu aktivitâte tįviōi un mâtîtes uzrâdîja pirmajâs 10 grûtniecîbas dienâs. Tįviōam visaugstâkâ aktivitâte tika konstatįta dzîvojot kopâ ar 12-25 dienas veciem mazuïiem un nevairojoðos dzîvnieku pâriem. Mâtîtįm visaugstâkâ aktivitâte tika konstatįta pirmajâs desmit laktâcijas dienâs, kâ arî dzîvojot kopâ ar diviem mazuïu metieniem un 12-25 dienas veciem mazuïiem. Lielu uzrâpðanâs reiþu skaitu varįja konstatįt grûtniecîbas laikâ neilgi pirms dzemdîbâm. Eksperimentos, kad bûrî pie dzîvniekiem tika ievietotas skaidas ar sveðu tįviōa smarþu, tika konstatįta ðîs aktivitâtes palielinâðanâs abu dzimumu îpatōiem(skat. 8. att.). Gan pirmâs, gan otrâs grupas dzîvniekiem lielâku uzrâpðanâs reiþu skaitu uzrâdîja tįviōð.
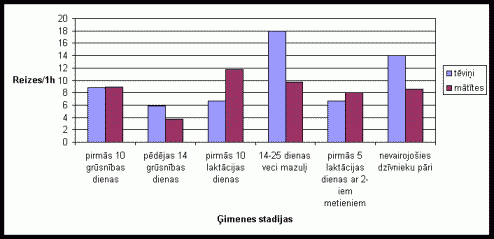
7. attįls Atraðanâs
skaits uz íieėeïiem vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem un
mâtîtįm atseviðíi, atkarîbâ no ėimenes stadijas.
(uz tekstu)
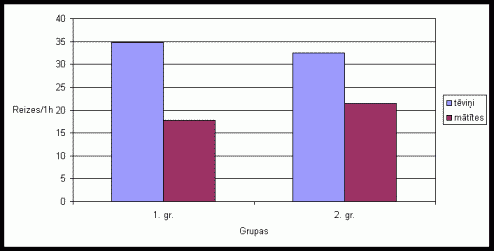
8. attįls Atraðanâs
skaits uz íieėeïiem vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem un
mâtîtįm, ja pirms eksperimenta bûrî tika ievietotas skaidasar sveða tįviōa
smarþu.
(uz tekstu)
Nazo-nazâlo un nazo-anâlo kontaktu skaits dzîvniekiem bija ïoti niecîgs gandrîz visu novįrojumu laikâ daþâdâs ėimenes stadijâs. Nazo-nazâlie kontakti tika konstatįti ïoti reti un parasti nepârsniedza 1-3 reizes. Nazo-anâlie kontakti tika konstatįti tikai daþas reizes visu novįrojumu laikâ. Nazo-nazâlo un nazo-anâlo kontaktu skaits strauji palielinâjâs ievietojot bûrî skaidas ar sveða tįviōa smarþu (skat. 9. att.). Gan pirmâs, gan otrâs grupas dzîvniekiem augstâku nazo-nazâlo kontaktu skaitu uzrâdîja mâtîtes, savukârt nazo-anâlo kontaktu skaitu- tįviōi. Seviðíi augstu aktivitâti uzrâdîja otrâs grupas tįviōi.
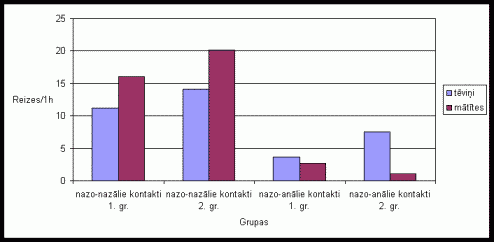
9. attįls Nazo-nazâlo
un nazo-anâlo kontaktu skaits vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem
un mâtîtįm, ja pirms eksperimenta bûrî tika ievietotas skaidas ar sveða
tįviōa smarþu.
(uz tekstu)
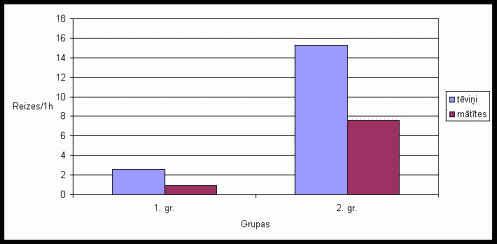
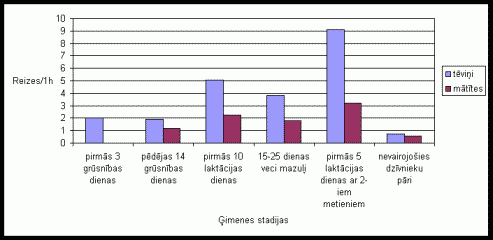
Aplûkojot pârskrįjienu skaitu vidįjos râdîtâjus varįja konstatįt, ka tįviōð parasti veic vairâk pârskrįjienu kâ mâtîte (skat. 10. att.). Pirmajâs trîs grûtniecîbas dienâs un dzîvojot kopâ ar 15-25 dienas veciem mazuïiem mâtîte vispâr neveica pârskrįjienus, arî tįviōð uzrâdîja samįrâ zemu pârskrįjienu aktivitâti pirmajâs trîs grûtniecîbas dienâs un vidįju aktivitâti dzîvojot kopâ ar 15-25 dienas veciem mazuïiem. Pįdįjâs 14 grûtniecîbas dienâs un dzîvojot kopâ ar diviem mazuïu metieniem, mâtîte vidįji veica vairâk pârskrįjienus. Interesanti ir tas, ka nevairojoðos dzîvnieku pâriem pârskrįjieni vispâr netika konstatįti. Eksperimentos, kad dzîvnieku bûrî tika ievietotas skaidas ar sveða tįviōa smarþu, bija novįrojamas samįrâ lielas atðíirîbas starp 1. un 2. grupas dzîvnieku aktivitâti (skat. 11. att.). 1. grupas dzîvniekiem gan tįviōi, gan mâtîtes veica samįrâ maz pârskrįjienu skaitu, savukârt otrâs grupas dzîvnieki izdarîja ïoti daudz pârskrįjienu, jo îpaði tįviōi.
10. attįls Pârskrįjienu
skaits vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem un mâtîtįm atseviðíi,
atkarîbâ no ėimenes stadijas.
(uz tekstu)
11. attįls Pârskrįjienu
skaits vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem un mâtîtįm, ja
pirms eksperimenta bûrî tika ievietotas skaidas ar
sveða tįviōa smarþu.
(uz tekstu)
Iegûtie rezultâti par uzmanîbas pozu skaitu un tajâs pavadîto laiku parâda, ka tįviōi ðo aktivitâtes veidu izrâda daudz bieþâk kâ mâtîtes un velta tam daudz ilgâku laiku(skat. 12. un 13. att.). Îpaði augstu aktivitâti tįviōi izrâdîja dzîvojot kopâ ar divu metienu mazuïiem, jo ðajos novįrojumos tika konstatįts ïoti liels uzmanîbas pozu skaits tįviōiem. Vįrtįjot uzmanîbas pozâs pavadîto laiku, varįja redzįt, ka visilgâko laiku ðajâs pozâs tįviōi pavadîja pirmajâs desmit laktâcijas dienâs, dzîvojot kopâ ar 15-25 dienas veciem mazuïiem un dzîvojot kopâ ar divu metienu mazuïiem. Mâtîte visaugstâko aktivitâti gan pozu skaita, gan tajâs pavadîtâ laika ziōâ izrâdîja dzîvojot kopâ ar divu metienu mazuïiem, bet pirmajâs trîs grûtniecîbas dienâs vispâr neizrâdîja ðo aktivitâti. Aptuveni vienâdi aktîvi bija nevairojoðies dzîvnieki.
12. attįls Uzmanîbas
pozu vidįjais laiks vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem un
mâtîtįm atseviðíi, atkarîbâ no ėimenes stadijas.
(uz tekstu)
13. attįls. Uzmanîbas
pozu skaits vienâ aktivitâtes ciklâ Íînas strupastu tįviōiem un mâtîtįm
atseviðíi, atkarîbâ no ėimenes stadijas.
(uz
tekstu)
Aplûkojot rezultâtus, kas iegûti reėistrįjot procentuâlos un vidįjos laikus, var redzįt, ka dzîvnieki įðanai velta aptuveni vienâdu laiku visâs ėimenes stadijâs (skat. 14. att. ). Tįviōð visilgâko laiku įðanai velta pirmajâs desmit laktâcijas dienâs, bet vismazâko laiku dzîvojot kopâ ar diviem mazuïu metieniem. Mâtîte visilgâko laiku įðanai velta pirmajâs desmit laktâcijas dienâs un dzîvojot kopâ ar 12 - 25 dienas veciem mazuïiem. Arî mâtîtes vismazâko laiku įðanai veltîjuðas, dzîvojot kopâ ar diviem mazuïu metieniem.
.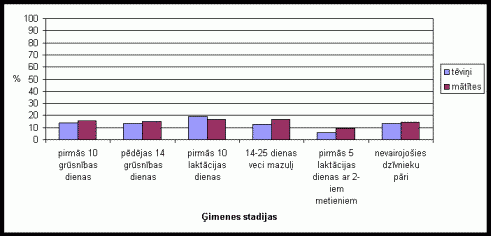
14. attįls. Įðanas
procentuâlais laiks (no kopįjâ aktivitâtes laika) Íînas strupastu tįviōiem
un mâtîtįm atseviðíi, atkarîbâ no ėimenes stadijas.
(uz tekstu)
Vienâ no eksperimentiem tika novįrota ïoti interesanta dzîvnieku uzvedîba, tâpįc ðos eksperimenta rezultâtus izdalîja atseviðíi. Ievietojot bûrî skaidas ar sveða tįviōa smarþu mâtîtes uzvedîba, pįc ðo skaidu apostîðanas, strauji izmainîjâs. Mâtîte sâka satraukti skraidît pa bûri, bieþi râpjoties uz íieėeïiem, bet ïoti reti urinįjot uz tiem. Mâtîtes aste bija nolaista. Pârskrįjieni un uzmanîbas pozas netika konstatįtas. Mâtîte bieþi tuvojoties savam tįviōam, râpâs tam virsû un veica t.s. uzsįdienus, kas raksturîgi tįviōam pâroðanas laikâ. Pįc kâda laika savs tįviōð tika izōemts no bûra un bûrî ievietoja sveðu tįviōu, kurð mįėinâja izbįgt no mâtîtes. Mâtîte neizrâdîja nekâdas agresijas pazîmes pret ðo tįviōu. Gluþi otrâdi, mâtîte mįėinâja ðim tįviōam tuvoties un veica uzsįdienus arî ðim tįviōam. Pįc tam sveðais tįviōð tika izōemts no bûra. Jâatzîmį, ka lîdz brîdim, kad mâtîte apoðōâja skaidas ar sveða tįviōa smarþu,seksuâla aktivitâte tai netika konstatįta.
Laktâcijas periodâ sâkumâ novįrotais zemais aktivitâtes stâvoklis apliecina to, ka tįviōi un mâtîtes pavada ïoti daudz laika migâ ar mazuïiem. Niecîgas kopîgi ârpus migas pavadîtais laiks, laktâcijas perioda sâkumâ liecina par to, ka kâds no vecâkiem gandrîz visu laiku atradâs migâ pie mazuïiem. Ðie rezultâti pierâda, ka Íînas strupastįm ir ïoti augsts vecâku tieðais ieguldîjums vecâku audzinâðanâ gan mâtîtįm, gan tįviōiem. To apstiprina arî ar nepârojoðajiem dzîvniekiem veiktie eksperimenti, kad to aktivitâte bija augsta. Ðie dati sakrît ar daudzu autoru iegûtajiem datiem, veicot eksperimentus ar monogâmo sugu grauzįjiem. (Dewsbury 1981; McGuire, Novak 1984;Solomon 1993; 1998). Interesanti atzîmįt, ka dzîvojot kopâ ar 12-25 dienas veciem mazuïiem ir novįrojams tįviōa ârpus migas aktivitâtes pieaugums. Mâtîtes aktivitâtes pieaugums nav novįrojams, taču var redzįt, ka dzîvnieki ðajâ laika ir pavadîjuði daudz lielâku laiku ârpus migas kopîgi. Tas uzskatâmi parâda, ka ðajâ laikâ dzîvnieki daudz mazâku laiku velta mazuïu aprûpei migâ. Îpaði augsta tįviōa aktivitâte tika novįrota dzîvojot kopâ ar divu metienu mazuïiem. Ðajâ laikâ ir novįrojams arî neliels mâtîtes aktivitâtes pieaugums un samįrâ augsta kopîgâ aktivitâte. Tas parâda to, ka daïu vecâku rûpju ðajâ laikâ uzōemas vecâkie mazuïi, kâ arî to, ka vecâkiem ðajâ laikâ ir nepiecieðams rûpįties par abiem mazuïu metieniem. Tįviōa paaugstinâtâ aktivitâte ðajâ ėimenes stadijâ ir konstatįta arî sugai Microtus ochrogaster (Solomon 1994). Tįviōa augstais vidįjais aktivitâtes lîmenis grûtniecîbas laikâ varįtu bût saistîts ar îpaðo aktivitâti, kas tika novįrota atseviðíâs dienâs grûtniecîbas sâkuma stadijâ, kâ arî grûtniecîbas beigâs, jo grûtniecîbas beigu posmâ, kâ arî grûtniecîbas sâkumâ mâtîte var atrasties pirmsestrusa vai estrusa stâvoklî. Jâatzîmį, ka atseviðíâs dienâs grûtniecîbas vidusposmâ tika novįrota ïoti augsta dzîvnieku aktivitâte. Veicot maksts uztriepju analîzi sugas Clethrionomys glareolus mâtîtįm, ir konstatįts, ka ðîs sugas mâtîtįm grûtniecîbas otrâs puses sâkumstadijâ ir raksturîgs stâvoklis, kas pielîdzinâms estrusa stâvoklim. Atseviðíos gadîjumos ir tikusi novįrota pat mâtîtes pâroðanâs ar tįviōu ðajâ grûtniecîbas stadijâ (Zaharovs, nepublicįti materiâli). Iespįjams, ðâdas hormonâlâs izmaiōas notiek arî Íînas strupastu mâtîtįm Tâ kâ ðîs sugas mâtîtįm estrusa laikâ ir raksturîga labprâtîga pâroðanâs ar sveðiem tįviōiem (Zakharov et al. 1998), tad ðî tįviōa aktivitâte, visticamâk ir saistîta ar partnera aizsardzîbu no pâroðanâs ar sveðu tįviōu. Ðâda partnera aizsargâðana ir ïoti raksturîga monogâmo grauzįju sugu tįviōiem (Carter et al. 1993; Insel et al. 1995). Mâtîtes augstâ aktivitâte pįdįjâs 14 grûtniecîbas dienâs, iespįjams, ir saistîti ar estrusa stâvokïa tuvoðanos, kura laikâ tâ var sâkt meklįt partneri (Zakharov et al. 1998), arî augsto kopîgo dzîvnieku aktivitâti ðajâ laikâ var izskaidrot ar mâtîtes estrusa stâvokïa tuvoðanos.
Gandrîz visâs ėimenes stadijâs tįviōi izrâda augstâku rakðanas aktivitâti kâ mâtîtes. Tas norâda, ka tįviōiem ir raksturîgs augsts ieguldîjums netieðajâs vecâku rûpįs par mazuïiem. Tįviōu augstâ rakðanas aktivitâte, dzîvojot kopâ 12 25 dienas veciem mazuïiem, varįtu bût skaidrojama ar atseviðío îpatōuîpaði augsto individuâlo aktivitâti, kâ arî ar to, ka mazuïi jau ir samįrâ pieauguði un lîdz ar to tįviōð var veltît laiku arî citâm aktivitâtįm, jo arî kopįjâ aktivitâte ðajâ laikâ pieaug. Dzîvojot brîvâ dabâ tįviōi ðo laiku, iespįjams, veltîtu savas fiziskâs formas uzlaboðanai. Pįdįjâs 14 grûtniecîbas dienâs un pirmajâs 10 laktâcijas dienâs novįrotâ augstâ tįviōu aktivitâte iespįjams ir skaidrojama ar tįviōu netieðajâm vecâku rûpįm. Ar to skaidrojama arî mâtîtes augstâ aktivitâte pirmajâs 10 laktâcijas dienâs. Augstâ rakðanas aktivitâte mâtîtįm, dzîvojot kopâ nevairojoðos dzîvnieku pâriem, visticamâk ir saistîta ar atseviðíu dzîvnieku augsto individuâlo aktivitâti kâ arî ar to, ka mâtîtes vairâk laika var veltît teritorijas ierîkoðanai. Savukârt nevairojoðies tįviōiem ðajâ laikâ nav attiecîga stimula, tâpįc to aktivitâte ir relatîvi zema.
Migas materiâla neðana, tâpat kâ rakðana ir pieskaitâma pie netieðajâm vecâku rûpįm. Tįviōu stabilâ migas materiâla neðanas aktivitâte visu ėimenes stadiju laikâ, liecina par augstajâm netieðajâm rûpįm par mazuïiem. Mâtîðu augstâ migas materiâla neðanas aktivitâte dzîvojot kopâ ar divu metienu mazuïiem ir saistîta ar to, ka, palielinoties kopâ dzîvojoðo dzîvnieku skaitam, jâpalielina arî migas izmįri. Nevairojoðos dzîvnieku zemâ migas materiâla neðanas aktivitâte pierâda to, ka migas materiâla neðana ir saistîta ar vecâku netieðajâm rûpįm un dzîvniekiem dzîvojot bez mazuïiem nav bijis attiecîga stimula ðîs aktivitâtes veikðanai. Tas apstiprinâjâs arî eksperimentos, kad dzîvnieku bûrî tika ievietots papildus migas materiâls. Ðajos eksperimentos aktivitâtes lielums migas materiâla neðanâ dzîvnieku grupâ, kas dzîvoja bez mazuïiem bija samįrâ augsts, tâpįc, ka ðajâ grupâ tika iekïauti visi dzîvnieku pâri, kas dzîvoja bez mazuïiem (lîdz ar to ðajâ grupâ atradâs arî grûsnas mâtîtes, tâpįc ðâdu dzîvnieku pâru aktivitâte, nesot migas materiâlu, bija samįrâ liela). Iegûtie rezultâti par rakðanu un migas materiâla neðanu, kas apliecina Íînas strupastu abu dzimumu îpatōu augsto ieguldîjumu netieðajâs vecâku rûpįs ir lîdzîgi vairâku pįtnieku rezultâtiem par monogâmo grauzįju vecâku uzvedîbu (Dewsbury 1981;McGuire, Novak 1984;Solomon 1993; 1998).
Urinâcija uz íieėeïiem var skaidrot kâ teritorijas iezîmįðanu, kas vairumâ gadîjumu tiek veikta, lai aizsargâtu teritoriju no savas sugas îpatōiem. Atseviðíos gadîjumos tįviōð urinâciju var veikt, lai aizsargâtu savu dzimumpartneri no pâroðanâs ar sveðiem tįviōiem. Augstâ tįviōa un mâtîtes urinįðanas aktivitâte dzîvojot kopâ ar 12 25dienas veciem mazuïiem ir skaidrojama ar teritorijas aizsardzîbu, bet tįviōiem arî ar mâtîtes aizsardzîbu, kas saistîta ar mâtîtes estrusa, pirmsestrusa vai estrusam lîdzîgo stâvokli. To apliecina arî dati, kas iegûti, veicot eksperimentus, kad dzîvnieku bûrî tika ievietotas skaidas ar sveða tįviōa smarþu. Raksturîgi, ka nevairojoðiem dzîvniekiem vairâk urinâciju veica mâtîtes, kas apliecina mâtîðu augsto teritorijas aizsardzîbas uzvedîbu. Otrâs grupas dzîvniekiem mâtîðu urinįðanas aktivitâte bija krietni zemâka, tas uzskatâmi parâda, ka mâtîtes teritorijas aizsardzîbas uzvedîbu bija pârmâkusi vairoðanâs uzvedîba un tâs, iespįjams, bija gatavas pâroties ar sveðo tįviōu. Ðîs grupas mâtîtes iezîmįjot teritorijas paaugstinâjumus tâdįjâdi varįja paziōot sveðajiem tįviōiem par savu gatavîbu pâroties. Tâpat atseviðíâm ðîs grupas mâtîtįm pâroðanâs gatavîba droði vien jau bija beigusies un, urinįjot uz íieėeïiem tâs izrâdîja teritorijas aizsardzîbas aktivitâti. Tįviōu lielais urinâciju skaits pirmâs eksperimenta grupas dzîvniekiem bija saistîts ar teritorijasaizsardzîbas aktivitâti, bet tâs straujais pieaugums otrâs grupas dzîvniekiem arî ar mâtîtes aizsardzîbu. Tįviōa augstâ urinįðanas aktivitâte, dzîvojot kopâ nevairojoðiem dzîvniekiem, visticamâk, ir skaidrojama ar teritorijas aizsardzîbu, kâ arî ar atseviðíu dzîvnieku augsto individuâlo aktivitâti. Lîdzîgi rezultâti tika iegûti veicot pįtîjumus ar sugas Phodopus campbelli dzîvniekiem (Wynne-Edwards, Lisk 1987).
Tįviōu un mâtîðu lielâ aktivitâti, râpjoties uz íieėeïiem, dzîvojot kopâ ar 12-25 dienas veciem mazuïiem ir skaidrojuma ar mâtîtes estrusa stâvokïa tuvoðanos, pįdįjâs ðîs ėimenes stadijas dienâs un iespįjamo estrusam lîdzîgo stâvokli ðîs stadijas vidusposmâ. Lîdz ar to palielinâs arî tįviōa aktivitâte, aizsargâjot mâtîti. Veicot eksperimentus, kad dzîvnieku bûrî tika ievietotas skaidas ar sveða tįviōa smarþu, konstatįtâ augstâ aktivitâte apstiprina ðiem dzîvniekiem raksturîgo augsto teritorijas aizsardzîbas aktivitâti. Zîmîgi, ka otrâs grupas mâtîtįm râpðanâs uz íieėeïiem bija bieþâk, nekâ urinįðana uz tiem, kas liecina par aizsardzîbas aktivitâtes samazinâðanos. Ðâdi, iespįjams, mâtîte izrâda aktivitâti, meklįjot dzimumpartneri. Lîdz ar to uzskatâmi parâdâs arî tįviōa aktivitâte aizsargâjot mâtîti. Tįviōa augstâ aktivitâte, dzîvojot kopâ nevairojoðiem dzîvniekiem, ir skaidrojama ar teritorijas aizsardzîbu, kâ arî ar atseviðíu dzîvnieku augsto individuâlo aktivitâti. Ðie izskaidrojumi, bieþi vien, neattiecâs uz tâm reizįm, kad dzîvnieki neurinįja uzrâpjoties uz íieėeïiem, jo tâdos gadîjumos dzîvnieku rîcîbu noteica izzinâðanas uzvedîba, kas raksturîga daudziem zîdîtâjiem.
Zemâ nazo-nazâlo kontaktu aktivitâte visâs ėimenes stadijâs liecina par to, ka ðâdâ ėimenes struktûrâ dzîvojoðiem dzîvniekiem nav nepiecieðama ðâda veida kontaktįðanâs, jo iespįja, ka to teritorijâ atradîsies sveðs îpatnis ir samįrâ neliela. Nazo-nazâlo kontaktu aktivitâtes paaugstinâðanâs, veicot eksperimentus, kuros dzîvnieku bûros tika ievietotas skaidas ar sveða tįviōa smarþu, liecina par to, ka ðajâ gadîjuma ir nepiecieðama ðâda kontaktįðanâs, lai konstatįtu vai teritorijâ nav potenciâlais sveðinieks un atpazîtu savus ėimenes locekïus. Pie ðîs aktivitâtes pieskaitâms arî augstais nazo-anâlo kontaktu skaits pirmâs grupas dzîvniekiem. Der atzîmįt, ka otrâs grupas dzîvniekiem novįrojamâ tįviōa augstâ un mâtîtes zemâ nazo-anâlo kontaktu aktivitâte skaidrojama ar mâtîtes aizsardzîbu, ko izrâda tįviōð (Zakharov et al. 1998). Tâdįjâdi tįviōð kontrolį vai mâtîte nav nonâkusi kontaktâ ar sveðo tįviōu un pârojusies ar to. Tâpat veicot eksperimentus ar citas monogâmas sugas Phodopus campbelli dzîvniekiem, tika konstatįts, ka atseviðíi sociâlâs uzvedîbas veidi ïoti mainâs atkarîbâ no estrâlâ cikla stadijas. Piemįram, apostîðanâs skaits dienâs, kad mâtîte bija gatava pâroties, bija divreiz lielâks kâ dienâs, kad mâtîte nebija gatava pâroties (Wynne-Edwards, Lisk 1987)
Lielais pârskrįjienu skaits, kas tika konstatįts tįviōiem, liecina par augstu tįviōu aktivitâti, aizsargâjot savu teritoriju. Pirmajâs trîs grûtniecîbas dienâs un pirmajâs 10 laktâcijas dienâs, konstatįtais augstais pârskrįjienu skaits ir izskaidrojams ar to, ka pirmajâs grûtniecîbas dienâs un pirmajâs pįcdzemdîbu dienâs, daþkârt, mâtîte var bût estrusam tuvâ stâvoklî. Savukârt pirmajâs grûsnîbas dienâs raksturîgâ mâtîtes neaktivitâte ðajâ uzvedîbas veidâ, liecina par tâs zemo agresivitâti un iespįjamo tieksmi pâroties. Pirmajâs 10 laktâcijas dienâs konstatįtais, augstais mâtîtes pârskrįjienu skaits ir attiecinâms uz ðîs stadijas beigu posmu, kad mâtîtei vairs nav tieksme pâroties, bet ðo uzvedîbas veidu nomâc mazuïu aprûpes uzvedîba, tâpįc mâtîtes teritorijas aizsardzîbas aktivitâte palielinâs. Neaktivitâte ðajâ uzvedîbas veidâ, kas tika konstatįta nevairojoðiem îpatōiem pierâda, ka dzîvniekiem, kuri neaudzina mazuïus un mâtîte atrodas seksuâli neaktîvâ stadijâ, ïoti pazeminâs aizsardzîbas aktivitâtes lîmenis, jo nav motivâcijas ðâdam aktivitâtes veidam. Lielas atðíirîbas dzîvnieku uzvedîbâ varįja konstatįt jau pieminįtajos eksperimentos. 1. grupas îpatōiem pârskrįjienu skaits bija ïoti zems, bet 2. grupas dzîvniekiem- izteikti augsts. No tâ var secinât, ka ðî aktivitâte lielâ mįrâ ir saistîta ar mâtîtes aizsardzîbu, ko spįcîgi izrâda otrâs grupas tįviōi. Tomįr, tâ kâ ðo aktivitâti izrâda arî mâtîtes, tas norâda arî par tâs nozîmîbu teritorijas aizsardzîbâ. No otrâs grupas mâtîtįm ðo aktivitâti, visticamâk izrâdîja mâtîtes, kuras vairs nebija seksuâlâ uzbudinâjuma stâvoklî.
Analizįjot rezultâtus, kas iegûti reėistrįjot uzmanîbas pozas, var secinât ka tįviōu augstâ aktivitâte, dzîvojot kopâ ar divu metienu mazuïiem ir izskaidrojama ar raksturîgo tįviōa vispârįjâs aktivitâtes palielinâðanos ðajâ ėimenes stadijâ. Arî mâtîtes augstâ aktivitâte ðajâ laikâ pierâda, ka ïoti lielas rûpes par jaunajiem mazuïiem izrâda vecâkie mazuïi, tâpįc mâtîte ilgâku laiku var pavadît, veicot citas aktivitâtes, tai skaitâ teritorijas aizsardzîbu. Tįviōa lielo un ilgstoðo uzmanîbas pozu skaitu, dzîvojot kopâ ar 15 25 dienas veciem mazuïiem un pirmajâs laktâcijas dienâs var izskaidrot ar to, ka grûtniecîbas beigu posmâ. laktâcijas sâkumâ, mâtîte var bût estrusa stâvoklî, savukârt grûtniecîbas vidus posmâ var parâdîties estrusa stâvoklim lîdzîga uzvedîba un hormonâlais stâvoklis. To pierâda arî mâtîtes zemâ aktivitâte ðajâ uzmanîbas veidâ. Tâdas paðas sakarîbas ir novįrojamas pirmajâs trîs grûsnîbas dienâs, kad mâtîte vįl var bût pįcestrusa stâvoklî.
Lai arî procentuâli pirmajâs 10 laktâcijas dienâs mâtîtes pavadîja salîdzinoði daudz laika, įdot ârpus migas, tomįr ōemot vįrâ îso laiku, ko tâs pavadîja ârpus migas ðajâ laikâ, var redzįt, ka vidįji įðana sastâdîja samįrâ maz laika. Lîdz ar to var secinât, ka lielâkâ įðanas aktivitâte mâtîtįm ir bijusi migâ, edot barîbu, ko tur sanesa tįviōi. Pįdįjâs 14 grûsnîbas dienâs konstatįtais paaugstinâtais mâtîtes įðanas lîmenis ir saistîts ar to, ka arî grûsnîbas laikâ mâtîtįm ir milzîgs enerėijas patįriōð, lîdz ar to mâtîtįm nepiecieðams uzōemt papildus barîbas vielas. Tįviōu zemais procentuâlais įðanas lîmenis, dzîvojot kopâ ar divu metienu mazuïiem, izskaidrojams ar ilgstoðo ârpus migas pavadîto laiku, lîdz ar to procentuâli įðanas laiks samazinâs, kaut arî redzams, ka vidįji tįviōi įðanai velta samįrâ ilgu laiku. Augstais procentuâlais įðanas lîmenis nevairojoðiem dzîvnieku pâriem ir saistîts ar zemo aktivitâti citos aktivitâtes veidos, lîdz ar to lielu daïu ârpus migas pavadîtâ laika dzîvnieki varįja veltît įðanai.
Atseviðíi izdalîtâ eksperimenta rezultâti, kura laikâ tika konstatįta mâtîtes seksuâlâs intereses izrâdîðana pret sveðu tįviōu, uzskatâmi parâdîjâs socio-seksuâlo attiecîbu pretrunîba monogâmo sugu dzîvniekiem, kas konstatįtas vairâkos pįtîjumos (Dewsbury 1981; Zakharov et al. 1998). Sociâlo monogâmiju, kâ sekmîgs mazuïu audzinâðanas veids nebût nenozîmį pilnîgu seksuâlo monogâmiju. Gluþi otrâdi Íînas strupastu mâtîtes ïoti labprât pârojas ar sveðiem tįviōiem (Zakharov et al. 1998). Tįviōð sakarâ ar iespįjamo mâtîtes neuzticîbu, izrâda pastiprinâtu dzimumpartnera apsargâðanu, laikâ, kad tâ spįjîga pâroties, gadîjumâ, ja teritorijâ konstatįtas pazîmes, kas liecina par sveða tįviōa iespįjamo klâtbûtni.
1. Abu dzimumu îpatōi izrâda augstas tieðâs vecâku rûpes.
2. Tįviōð izrâda ne mazâkas netieðâs vecâku rûpes kâ mâtîte.
3. Abu dzimumu îpatōi izrâda uzvedîbu, kas saistîta ar teritorijas aizsardzîbu, tįviōð izrâda arî ïoti aktîvu partnera aizsardzîbu no pâroðanâs ar sveðiem tįviōiem.
4. Neskatoties uz Íînas strupastįm raksturîgo sociâlo monogâmiju un abu partneru augsto ieguldîjumu mazuïu aprûpį, tâm tomįr nav raksturîga absolûtâ seksuâlâ monogâmija.
 |
||||||||||||
| LU | LU BF | LDF | Ezeri | Sugu enciklopÄdija |
BioloÄĢiskÄ daudzveidÄŦba |
Latvijas daba | Piekrastes biotopi |
Malokologu biedrÄŦba |
BotÄnikas biedrÄŦba |
MeklÄ | Raksti mums |