1.1.2. Limnoplanktona īpatnības
2.1. Zooplanktona pētīšanas metodes
3. Kaņiera ezera Cladocera fauna 2001.gada vasarā
1.attēls
2.attēls
3.attēls
4.attēls
5.attēls
6.attēls
7.attēls
8.attēls
Kladoceras (Cladocera) ir vēžveidīgo apakštipa (Crustacea) žaunkāju klases (Branchiopoda) kārta.
Kladoceras ir saldūdeņos un jūrās plaši izplatīta vēžveidīgo grupa ar lielu ekoloģisku un praktisku nozīmi.
Cladocera
fauna ir viens no būtiskākajiem trofiskā tīkla un bioloģiskās daudzveidības
elementiem.
Darba mērķis – analizēt Kaņiera ezera Cladocera faunu.
Darba uzdevumi:
Planktons (no grieķu ”planktos” – klejojošs) ir mazu organismu kopa, kas apdzīvo kontinentālus un jūras ūdeņus (БСЭ, 1975), bez vai ar ierobežotām spējām pārvietoties (Wetzel,1995). Aktīvās pārvietošanas gadījumā mazo ātrumu dēļ tas nespēj pretoties ūdens straumēm (Константинов,1986). Planktons ir suspendēts ūdenī un lielākoties tiek izplatīts ar turbulenci un citām ūdens kustībām (Wetzel,1995).
Planktona sastāvā ietilpst kā augi – fitoplanktons tā arī bakterioplanktons un dzīvnieki – zooplanktons (БСЭ, 1975), kas parasti ir blīvāks par ūdeni un grimst gravitācijas ietekmē (Wetzel, 1995).
Daudzi planktona dzīvnieki veic regulāras vertikālas migrācijas ar daudzu metru amplitūdu. Migrācijas sekmē barības vielu resursu pārnesi no virsējiem, ar tām bagātiem, slāņiem dziļumā.
Planktons tieši vai caur starpposmiem kalpo kā barības bāze daudziem cilvēkam vērtīgiem dzīvniekiem (zivīm, jūrās – kalmāriem, vaļiem u.c.).
Planktonorganismu izmēri svārstās no dažiem mikrometriem līdz dažiem metriem. Līdz ar to parasti izšķir :
ultraplanktonu (baktērijas; organismu izmēri nepārsniedz dažus mikrometrus, zemākā robeža ir aiz redzamības sliekšņa) (Яшнов, 1969);
nannoplanktonu (baktērijas, sīkākas vienšūnas aļģes);
mikroplanktonu (lielākā daļa aļģu, vienšūņi, virpotāji, daudzi kāpuri)
mezoplanktonu ( kopepodi, kladoceras un citi dzīvnieki, mazāki par 1cm)
makroplanktonu (daudzas mizīdas, garneles, medūzas u.c. salīdzinoši lieli dzīvnieki)
megaplanktonu, pie kura pieskaita nedaudzus lielākus planktondzīvniekus (jūras medūza cianeja (diametrs līdz 2m, taustekļi līdz 30m), pirosomu kolonijas līdz 30m garumā u.c.), taču šo grupu izmēru robežas nav viennozīmīgi pieņemtas (БСЭ,1975, Константинов,1986).
Saldūdeņos izšķir ezeru planktonu (limnoplanktonu) un upju planktonu (potamoplanktonu) ( БСЭ, 1975).
1.1.2. Limnoplanktona īpatnības ![]()
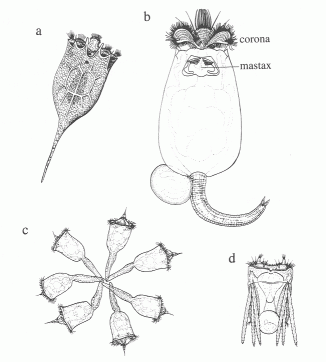
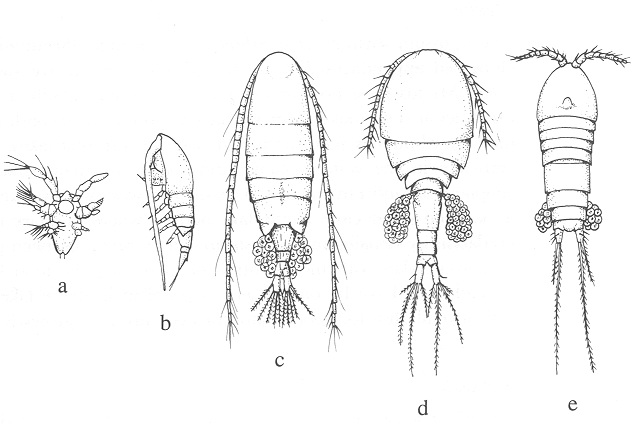
Ezera zooplanktona galvenās sastāvdaļas veido mikrozooplanktons, ko sastāda vienšūņi (Protozoa) un virpotāji (Rotatoria) (1.attēls), un mezozooplanktoniski posmkāji (Crustacea), kuriem pieskaitāmas kladoceras (Cladocera) un airkājvēži (Copepoda, Calanoida) (2.attēls). Dažos ezeros makrozooplanktons, tāds kā Chaoborus kāpuri un Mysis sp. ir lielā daudzumā.
Zooplanktona izdalītās pārstrādātās barības vielas var būt galvenais slāpekļa (N) un fosfora (P) avots termālās stratifikācijas laikā.
Vairums zooplanktona ir aptuveni 0.5-3 mm gari, lielākā daļa barojas ar aļģēm vai baktēriju sakopojumiem ar diametru 1-15 mm. Copepoda un Rotatoria var aprīt citu zooplanktonu.
Liels zooplanktona blīvums svārstās no aptuveni 500 eks/litrā eitrofajos ezeros līdz mazāk par 1 eks/litrā oligotrofos ūdeņos (Horne, Goldman, 1994).
Pie zooplanktona adaptācijām dzīvei ūdenī pieskaitāmi
Zooplanktons bieži ir caurspīdīgs, kas samazina zivju plēsonību.
Dažiem ir dzeloņi, lai nodrošinātu aizsardzību pret bezmugurkaulnieku plēsoņām.
Ātra partenoģenētiska vairošanās samazina plēsonības ietekmi un ļauj dažiem planktoniskiem dzīvniekiem izmantot ātri augošu aļģu skaita uzplaukumu (Horne, Goldman, 1994).
Zooplanktona sastāvs visa gada laikā nav vienāds (Sloka,1961). Cladocera un Rotatoria ir lielākā daudzumā vasarā, kad tiem ir vairāk barības. Copepoda un Mysis ir vairāk daudzgadīgi, ar lielu pārziemojošu populāciju. Daudziem Cladocera un Rotatoria ir miera stāvokļi un izturīgas olas, kas pārziemo nogulumos vai tiek ar vēju vai putniem pārnesti uz jaunām dzīvesvietām.
Zooplanktons ir galvenā saite starp mazu fitoplanktonu un lielākiem plēsējiem, pirmkārt, zivju mazuļiem.
Lielākā daļa zooplanktona barojas satverot lielāku upuri vai noslaukot aļģes, baktērijas no ūdens, un koncentrē šo barību gremošanai, filtrējot cauri ar sariņiem klātām mutes ekstremitātēm.
Dzīvnieku planktona izplatību un daudzumu ierobežo barības pieejamība un plēsonība.
Posmkāju zooplanktons (Crustacea) ir mazi, puscaurspīdīgi organismi, kas, kopā ar pelaģiskiem vienšūņiem (Protozoa) un virpotājiem (Rotatoria) sastāda lielāko daļu no saldūdeņu dzīvnieku planktona ( Horne, Goldman, 1994).
1.2. Cladocera kā ezeru zooplanktona funkcionālā grupa ![]()
Šobrīd pasaulē ir sastopamas gandrīz 400 kladoceru sugas, kuras tiek dalītas 4 apakškārtās pretēji kādreizējam iedalījumam 4 kārtās (Dosson, Frey,1991). Pēdējie molekulārie pētījumi liecina, ka Cladocera ir monofilētiska grupa (Crease, Taylor, 1998). Latvijā konstatētas 11 dzimtas ar vairāk nekā 80 sugām (Sloka, 1981).
Cladocera sistemātika (Crease, Taylor, 1998)
Klase Branchiopoda Latreille, 1817
Apakšklase Diplostraca Gerstaecker, 1866
Kārta Cladocera Latreille, 1829
Apakškārta Anomopoda
Dzimtas: Daphniidae Straus, 1820
Moinidae Goulden, 1968
Bosminiidae Baird, 1846
Macrothricidae Norman & Brady, 1867
Chydoridae Stebbing, 1902
Apakškārta Ctenopoda
Dzimtas: Sididae Baird, 1850
Holopedidae Sars,1865
Apakškārta Onychopoda
Dzimtas Polyphemidae Baird, 1845
Cercopagidae Mordukhai-Boltovskoi, 1966
Pogonidae Mordukhai-Boltovskoi, 1968
Apakškārta Haplopoda
Dzimta Leptodoridae Lilljeborg, 1861
1.2.2.Cladocera bioloģija un ekoloģija ![]()
Kladoceras ir mazi, parasti caurspīdīgi vēžveidīgie, kas parasti tiek sauktas par “ūdens blusām” to formas un lecošo kustību dēļ (Brönmark, Hansson, 2001). Kladoceras ir svarīga biocenožu sastāvdaļa. Spēdamas labvēlīgos apstākļos strauji savairoties lielā skaitā, tās veido nozīmīgu posmu barošanās ķēdēs. No vienas puses, kladoceras regulē baktēriju un detrīta daudzumu, no otras – ir nozīmīgs komponents zivju mazuļu, planktonēdāju zivju un daudzu citu ūdens dzīvnieku barībā, tāpēc arī ūdenstilpju bioloģisko produktivitāti bieži raksturo pēc Cladocera sugu sastāva un biomasas. Bieži tās izmanto arī kā bioindikatorus ūdens piesārņojuma raksturošanai. Visos šajos gadījumos runa ir galvenokārt par mātītēm, jo tēviņi parādās tikai īsu brīdi, kamēr dzīves cikla lielākajā daļā mātītes vairojas partenoģenētiski.
Mātīšu ķermeņa garums svārstās no 0.2-10 mm (Sloka, 1981).
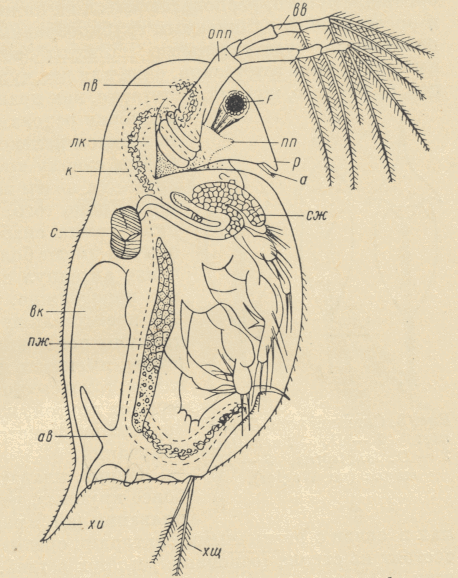
Kladoceru ķermenis sastāv no vairāk vai mazāk atdalītām galvas, krūtīm un postabdomena. Kopējais segmentu skaits nav noteikts, jo ķermeņa pakaļējais gals nav segmentēts, un varētu svārstīties no10-15 (3.attēls).
Ķermenis ir klāts ar vairāk vai mazāk caurspīdīgu hitīna kutikulu, kas ir veidojusies no hipodermiskā epitēlija slāņa zem tās. Lielākai kladoceru daļai hitīna kutikula veido divvāku čaulu, kas piesedz ķermeni ar ekstremitātēm. Čaulas virsmas struktūra ir daudzveidīga, parasti tā ir retikulēta. Bieži uz čauliņas veidojas krokas – ķīļi. Vāciņu dorsālā daļa dažreiz ir izstiepta īsākā vai garākā dzeloņveidīgā izaugumā.
Dažādu formu galva ir virzīta uz leju, dažreiz ļoti maza (Ceriodaphnia) vai ļoti liela (Bythothrephes).Uz galvas hitīna kutikula veido vairogu, kura mugurējā daļa savienojas ar pārējo ķermeni ar vai bez skaidras robežas. Galvas vairoga priekšējā mala bieži ir knābjveida izstiepta un veido t.s. rostrumu. Daudzām kladocerām (galvenokārt, tām, kurām ķermenis ir stipri saspiests no sāniem) uz galvas mugurējās malas ir ķīlis, visbiežāk gluds, dažreiz ar zobiņiem (Daphnia carinata), dažreiz paplašinās par pakavveida plāksnīti (D. atkinsoni). Daudzām kladocerām ir laterāls galvas ķīlis, kas sākas virs acs un stiepjas pie peldantenu pamata (Мануйлова, 1964). Galvas dorsālajā pusē var būt galvas poras, to forma ir dažāda. Lielās sauc par makroporām, mazās – par mikroporām. Sugu vairumam ir liela melna acs, dažām arī (parasti par to mazāka) nauplijacs (Sloka, 1981). Uz galvas ir 5 piedevu pāri – antenas, peldantenas,mandibulas, maksilas, maksilulas.
Antenulas ir maņu orgāni, klāti ar jutīgām papillām (estetām) distālajā galā un sariņiem. Antenulu uzbūve atšķiras tēviņiem un mātītēm.
Peldantenas ir lielākas par antenulām, novietotas galvas sānos (Sloka, 1961). Peldantenas ir divzarotas, vienīgi Holopedium mātītēm tās ir nezarotas (Sloka, 1981).
Krūts piedevas ir 4-6 kāju pāri, kas ir pilnīgi zaudējušas kustības funkciju un pārvērtušies par sarežģītu aparātu barības iegūšanai. Lielākoties kladocerām kājas veido filtrētājaparātu, biezi klātu ar sariņiem (Мануйлова, 1964). Dažām sugām kājas veic līdz 500 kustību minūtē (Догель, 1974). Kājas ir divzarotas, to ārējo zaru sauc par ekso-, iekšējo – par endopodītu. Krūtīm seko vēders bez ekstremitātēm. To dorsālajā pusē ir divi gari abdomināli sari un var būt abdomināli izaugumi. Vēdera distālais gals sašaurinās un veido postabdomenu (kaudu, abreptoru), kas noliecās uz leju un beidzas ar diviem nagiem vai garu kaudālo piedevu. Postabdomena dorsālās malas katrā pusē parasti izvietojas dzelkņi vai sari (Sloka, 1981).
Anālā atverevisbiežāk ir postabdomena augšējā malā.
Iekšējais skelets sastāv no membrānu un stiegru sistēmas, kas kalpo galvenokārt muskuļu piestiprināšanai un kopā ar tiem piedalās asins cirkulācijas regulācijā.
Muskulatūra ir bez metamērijas. Ir gareniski, šķērseniski un dorsoventrāli muskuļu kūlīši.
Nervu sistēma sastāv no divām ventrālās nervu ķēdītes stiegrām ar pāra ganglijiem. Plēsīgām kladocerām daļa no pāru ganglijiem ir saplūduši. Bez tiem, ir arī pāra virsrīkles ganglijs un rīkles komisūras.
No maņu orgāniem vislabāk attīstīti redzes orgāni –saliktā nepāra fasetacs un nauplijacs, kas var būt reducēta plēsīgām formām.
Ķīmiskās analīzes orgāni ir estetas uz antenulām. Estetu skaits dzimtā ir patstāvīgs (Polyphemidae – 5, Sididae, Daphniidae, Bosminiidae –9) un parasti ir lielāks tēviņiem. Taustes orgāni ir jutīgi sariņi, taustes sariņš antenulu priekšējā malā, viciņa uz tēviņu antenulām, sariņi peldantenu pamatā, astes sariņi.
Gremošanas sistēma. Gremošanas kanāls dalās trijās aptuveni vienādās pēc garuma daļās- barības vadā (priekšējā zarna), viduszarnā un galazarnā.
Barības vada dobumu izklāj kutikula, sieniņas ir ar iekšējo gredzenisko un ar ārējo garenisko muskulatūru.
Viduszarna lielākoties ir taisna, bet Chidoridae un dažiem Macrothricidae cilpveidīgi izlocīta. Daphniidae priekšējā viduszarnas daļa veido pāra aklus izaugumus, t.s. aknu izaugumus.
Īsa galazarna, nedaudz paplašināta pie anālās atveres, ir izklāta ar hitīna kutikulu, ir bez gareniskās muskulatūras, bet ar spēcīgiem muskuļiem-paplašinātājiem.
Asinsrites sistēma ir vāji attīstīta. Ir maisveida vai vārpstveida sirds, aizmugurē noslēgta un ar atveri priekšpusē. Abos sānos ir ostiju pāris. Sirds saraušanās notiek ātri (līdz 250 reižu minūtē). Dobuma šķidrums (asinis) ir bezkrāsains vai vāji iekrāsots (sarkans vai dzeltens), tajā ietilpst sīkas, 7-8 mm diametrā amēbveida šūnas. Asinsvadi nav attīstīti. Asinis no sirds nokļūst starporgānu telpā, asins pārvietošana pa ķermeni noris ar iekšējā skeleta membrānu un starpsienu palīdzību.
Elpošanas sistēmu lielākai kladoceru daļai veido žaunu maisiņi epipodīti, kas piepildīti ar tajos cirkulējošām asinīm. Difūzā elpošana notiek caur galazarnas sieniņām un īpašu pakauša orgānu, kas ir sastopams jauniem īpatņiem.
Izvadorgānu sistēma sastāv no maksilāro dziedzeru pāra, kas ir rudimentālo maksilu piedevas un atveras pie to pamata. Metanefrīdiji sastāv no celomiskā maisiņa nefrostoma bez skropstiņām, kas pāriet garā nefridiālā kanālā. Kanāls met trīs cilpas un veido nelielu savācējpūslīti, kas atveras pie maksilu pamata. Nefridiālā kanāla saturs (metabolisma galaprodukti) izdalās ekskrēta pilienu veidā vai tiek izsviests ātros grūdienos.
Dzimumsistēmu tēviņiem un mātītēm veido līdzīgas uzbūves dzimumdziedzeri.
Olnīcu pāra caurulītes stiepjas gar gremošanas trakta malām un atveras perējamā kamerā. Olnīcā ir divi – embrionālais un augšanas- slāņi.
Arī sēklinieki cauruļu veidā novietojas gar gremošanas kanālu. Visbiežāk sēklvadu atveru vietā veidoja nelieli pauguriņi – papillas. Spermatozoīdi bez astes piedevas var būt ieapaļi vai dzeloņainās plāksnītes veidā.
Apaugļošanāsvisbiežāk norisinās brīdī, kad tēviņš piestiprinās pie mātītes čauliņas pakaļējās malas. Mātītes satveršanai kalpo nadziņi uz tēviņa pirmā kāju pāra, arī antenulu viciņa. Apaugļošanas laikā sperma tiek izlaista olvadu atveru tuvumā (Мануйлова, 1964).
Dzīves cikls. Kladoceras ir spējīgas attīstīties partenoģenētiski. Telpa starp karapaksu (čauliņu) un ķermeni kalpo mātītēm kā perējamā kamera. Te olas attīstās par embrijiem, kas laika gaitā pieņem pieaugušo īpatņu formu un vēlāk tiek izlaisti ūdenī. Partenoģenētiskā reprodukcija turpinās līdz nelabvēlīgo laika apstākļu iestāšanās perioda, tad dažas olas pārvēršās par tēviņiem, pārējās – par haploīdām olām, kam nepieciešama apaugļošana. Tēviņu parādīšanos ietekmē liels populācijas blīvums vai strauja barības vielu samazināšanās. Pēc apaugļošanās perējamās kameras sieniņas uzbiezinās, veidojot ziemas olas (ilgolas), kas ir viegli pamanāmas kā tumši ovāli veidojumi. Ilgolas var paciest bargus apstākļus, tādus kā iesaldēšana vai izžūšana, tās var tikt tālu izplatītas (piemēram, ar putnu palīdzību). Labvēlīgos apstākļos no ziemas olām izšķiļas jaunas partenoģenētiskas mātītes, kas ātri savairojas (Brönmark, Hansson, 2001).
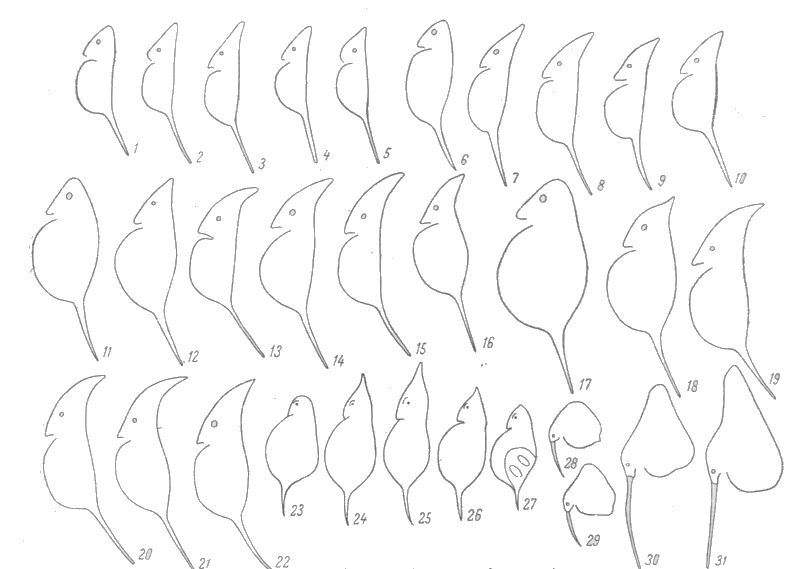
Dažam kladocerām (Bosmina, Daphnia) dažādu sezonu paaudzēm čaulas forma un izmēri stipri atšķirīgi, t.i., tām vērojama ciklomorfoze (4.attēls) (Sloka, 1981).
Barošanās. Daudzas kladoceras, ieskaitot Daphnia un Bosmina, ir pelaģiski filtrētāji, kas pārtiek no aļģēm un noteiktām baktērijām. Daphnia ir ģenerālisti – filtrētāji ar plašu barības spektru, to zarnu saturs var kalpot par labu kvalitatīvu apkārtējo ūdeņu aļģu un baktēriju sugu rādītāju. Citas kladoceras, piemēram, Chydorus, ir bentiskas, tipiskākajā gadījumā barojas, rāpojot gar virsmām, noskrāpējot un filtrējot barības daļiņas. Lielas kladocerasLeptodorus, savukārt, ir vairāk plēsīgas, ķer lielus upurus, tādus kā Rotatoria, Ciliata un pat Copepoda (Brönmark, Hansson, 2001).
2.1. Zooplanktona pētīšanas metodes
![]()
Zooplanktona paraugu ņemšanai izmantojamā aparatūra un metodes ir atšķirīgas dažāda tipa, lieluma un dziļuma ūdenstilpēs (Zandmane, 1995).
Planktoniskus dzīvniekus (vienšūņus, virpotājus, kladoceras, airkājus, daudzu bentisku dzīvnieku kāpurus) ievāc ar planktona tīkliņu (5.attēls).
Tīkliņa gredzens vākšanas laikā jātur zem ūdens. Kvantitatīviem pētījumiem caur Apšteina tīkliņu filtrē noteiktu ūdens daudzumu (Sloka, 1971), sakoncentrē līdz 200 ml apjomam. katrā paraugu ņemšanas vietā ar spaini tīkliņā ielej 100 l ūdens (Zandmane, 1995), tīkliņa konusā sakrājas atfiltrētais paraugs, kuru tālāk pārlej plastmasas pudelē vai nu atgriežot īpašu atgriezni, vai atsienot tīkliņa galu (Sloka, 1971).
Ievākto paraugu fiksē ar 40% formalīnu līdz 4% koncentrācijai paraugā (1 daļa formalīna uz 9 daļām parauga).
Mikroplanktona ievākšanai izmanto tīkliņu Nr. 64-77, mezoplanktona ievākšanai – Nr. 38-64. nannoplanktoma ievākšanai izmanto citas metodes.
Tīkliņa numurs norāda tīkliņa izgatavošanai izmantotā gāzauduma vai kaprona sieta blīvumu, t.i. acu skaits uz 1 cm2 sieta (Zandmane, 1995).
2.2. Pētīto paraugu ievākšanas
metode ![]()
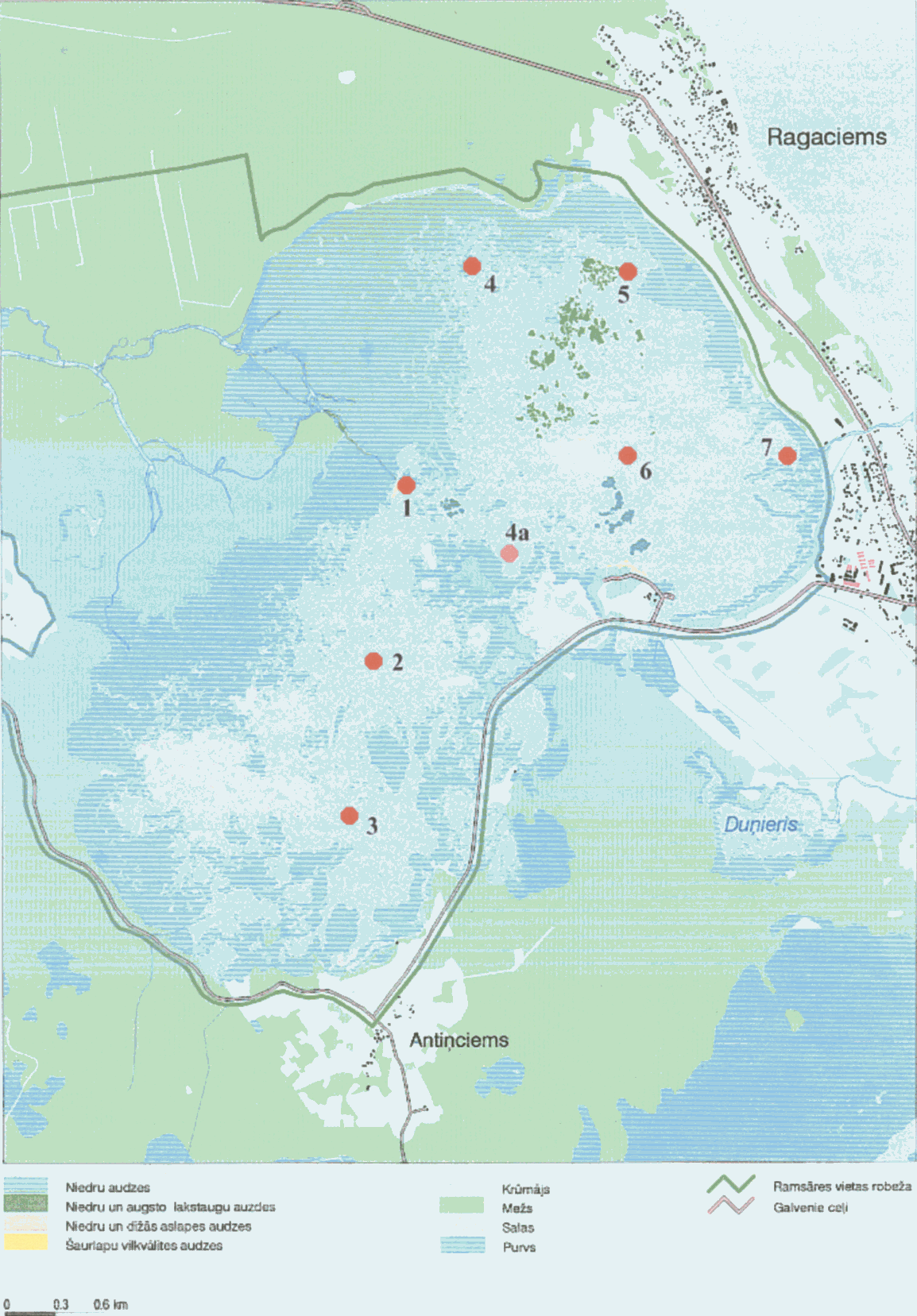
Pētītie paraugi tika ievākti 31.05.2001, 7.08.2001, 9.08.2001 un 12.10.2001 Kaņiera ezera septiņās stacijās (6.attēls).
Ievākšanai 100 l ūdens tika filtrēts caur standarta planktontīkliņu, paraugi fiksēti ar 40% formalīnu līdz 4%formalīna koncentrācijai paraugā.
Paraugi tika analizēti kvalitatīvi, sugu noteikšanai izmantoti noteicēji:
Paraugu pētīšanai izmantots mikroskops Amplival (Carl Zeiss Jena), 16x10 (160) palielinājumā.
2.3. Kaņiera ezera hidrobioloģiskais raksturojums ![]()
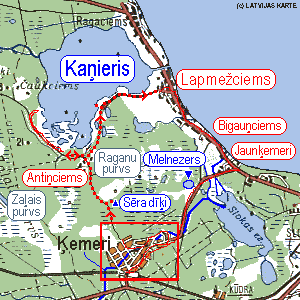
Kaņiera ezers (Kaņieris) atrodas Tukuma rajona austrumu daļā starp Ragaciemu, Lapmežciemu, Antiņciemu un Čaukciemu (7.attēls,8. attēls) (Smaļinskis, 1997), Lapmežciema pagastā (www.fishing.lv), 2.1m vjl (Lūmane, 1995).
Kaņieris ir piekrastes lagūnas tipa ezers, kas ir izveidojies no Litorīnās jūras lagūnas. Ar ūdens līmeņa pazemināšanos pamazām izzuda tā saikne ar jūru.
Kaņieris ir Latvijas 13. lielākais ezers (Smaļinskis, 1997). Ezera maksimālais garums 5.2 km, platums – 3.6 km, krasta līnijas garums 21.8 km (www.ezeri.lv), pašreizējā platība 11.28 km2, ar salām – 11.44 km2 (Smaļinskis, 1997). Ezera maksimālais dziļums 1.8 m, vidējais dziļums – 0.6 m, tilpums 6.68 mlj. m3( www.ezeri.lv).
No rietumiem ezerā ieplūst lielākā upe Slocene (Pulkaine), tās vidējais caurplūdums 2.44 m3/s (Lūmane, 1995), mazāka Medupīte, vairāki grāvji (www.ezeri.lv). Pašlaik Kaņiera ūdeņi uz Baltijas jūru plūst pa 0.7 km garu Starpiņupīti. Ziemeļos un ziemeļaustrumos Kaņieri no Baltijas jūras atdala apmēram 1 km plata sauszemes josla (Smaļinskis, 1997). Sateces baseins uz 1972. gadu bija 357 km2 (www.ezeri.lv).
Ezera ūdens līmenis un platība vairākkārt tika mainīti. 19.gs. tā platība bija ~20 km2 (Smaļinskis, 1997), līmenis – 2-2.3 m vjl. 20.gs sākumā pēc Starpiņupītes izrakšanas ūdens līmenis ir krities un ezera platība ir samazinājusies, tomēr plāns jaunas lauksaimniecībā izmantojamas zemes iegūšanai nav īstenojies, un 40.gadu sākumā, uzbūvējot dambi, ūdens līmenis atkal tika paaugstināts. Kaņiera ūdens līmeņa pārmaiņas radīja negatīvas sekas: ezers sadalījās vairākās daļās, pārpurvojās lielas platības, samazinājās lielā putnu un zivju daudzveidība. Tas varēja nelabvēlīgi ietekmēt Ķemeru kūrorta turpmāko attīstību, tāpēc 1964.g. atjaunoja ezera agrāko līmeni, ierīkojot uz Starpiņupītes divas slūžas. Līdztekus ezerā tika izveidotas 5 mākslīgas salas, lai piesaistītu ligzdojošos ūdensputnus (www.jurmala.lv).
Kopējais salu skaits ezerā ir 14, to platība 15.9 ha (www.ezeri.lv), lielākās ir Krievu un Riekstu salas (Smaļinskis, 1997).
Ezera grunts galvenokārt dūņaina (www.fishing.lv), vietām dūņas līdz 6.5 m, vietām akmeņaina, austrumu daļā gruntis dolomīta, ar nelielu dūņu slānīti (www.ezeri.lv), piekraste smilšaina (www.fishing.lv). Dūņas izmanto balneoloģijā (Lūmane, 1995).
Austrumu krasti stingri, pieejami, zemi;
D, DR –purvaini, lēzeni;
R- staigni, grūti pieejami;
Z – dūņaini (www.ezeri.lv).
Krasta līniju dienvidos un austrumos izrobo līči un pussalas (Lūmane, 1995).
Apkārtni veido
A- krūmājs, ceļš starp Lapmežciemu un Antiņciemu;
R- purvs bez piebraucamiem ceļiem;
ZA- Lapmežciems, ir piebraucami ceļi un ceļš Jūrmala- Engure;
Z- Ragaciems, ceļš Jūrmala- Engure.
Peldvietas un makšķernieku bāze ierīkotas ZA krastā.
Augsnes daudzveidības un hidroloģisko īpatnību dēļ ir liela augu dažādība. Kaņiera krastos sastopamas daudzas retas sugas: purva dievkrēsliņš (Euphorbia palustris), Pallasa sausserdis (Lonicera pallasii), tumšzilā drudzene (Genciana pneumonante), dzeltenā dzegužkurpīte (Cypripedium calceolus), Lezeļa lipāre (Liparis loeselii), mežloki (Allium ursinum) u.c. Ezera krastos aug tādi atlantiska laika relikti kā dižā aslape (Cladium mariscus), rūsganā melncere (Schoenus ferrugineus) un parastā purvmirte (Myrica gale). Virsūdens aizaugumu veido galvenokārt niedres (Phragmites australis), grunts lielāko daļu sedz mieturaļģu (Chara) audzes.
Kaņiera ezerā ir reģistrētas vairāk kā 20 zivju sugas, daudzas no tām veido stabilas populācijas. Visvairāk pārstāvētas ir līdakas (Esox lucius), plauži (Abramis brama), raudas (Rutilus rutilus), līņi (Tinca tinca), asari (Perca fluviatilis), karūsas (Carassius sp.) (Smaļinskis, 1997). Katru gadu zivju krājumi tiek papildināti ar līdaku mazuļiem (www.fishing.lv).
Lielākā daļa Latvijas abinieku un rāpuļu sugu izmanto ezeru un tā apkārtni kā dzīves, barošanās un vairošanās vietu.
Kaņieris ir viens no Latvijas ar putniem bagātākiem ezeriem. Nevelti vārda “Kaņieris” izcelsmi skaidro ar lībiešu “Kan järve”, kas tulkojumā nozīmē “zosu ezers”. Kopumā ezerā un tā apkārtnē ir reģistrētas 237 putnu sugas, vismaz vienreiz ligzdojot no tām ir novērotas 177. No sugām, kuras ir ierakstītas Latvijas Sarkanajā grāmatā, ezeru krastos un tā apkārtnē ligzdo 48 sugas (62%): lielais dumpis (Botaurus stellaris), mazais dumpis (Ixobrychus minutus), melnais stārķis (Ciconia nigra), paugurknābja gulbis (Cygnus olor), meža zoss (Anser anser), niedru lija (Circus aeroginosus), jūras ērglis (Haliaetus albicilla), grieze (Crex crex), gugatnis (Phylomachus pugnax), ūpis (Bubo bubo) u.c. Kuplā skaitā pārstāvētas dažādas pīļu sugas – maža pīle (Anas platyrhynchos), brūnkaklis (Aythia ferina) u.c., tomēr skaitliski vairāk ir lielais ķīris (Larus ridibundus). Kā spalvu maiņas vietu ezeru izmanto ~2000 pīles, kā atpūtas un barošanās vietu – gājputni.
Jau kopš 1939. g. Kaņiera ezers pieder pie Ķemeru sēravotu dabas rezervāta. Lai aizsargātu ligzdojošus un gājputnus, 1964. g. tika izveidota Kaņiera ezera 1857 ha liela zooloģiskās aizsardzības teritorija. Kopš 1977. g. cilvēka klātbūtne šajā dabas rezervātā (D un R daļā) bezledus periodā ir aizliegta, bet pārējā aizsargājamās teritorijas daļā (759 ha) nedrīkst uzturēties pavasara migrācijas un ligzdošanas laikā. 1989. gadā ezers tika iekļauts starptautiskās putnu aizsardzības biedrības (Birdlife International) Eiropas nozīmīgāko putnu vietu reģistrā. Kopš 1995.g. Kaņiera ezers ir iekļauts Rāmsāres konvencijas vietu reģistrā kā dabas aizsardzības objekts ar starptautisku nozīmi.
Kaņiera ezers, krasti, purvi un mitrie meži pieder pie Ķemeru nacionālā parka teritorijas (Smaļinskis, 1997), kas tika nodibināts 1997. gadā (www.jurmala.lv)
Nopietnākie draudi Kaņierim ir lauksaimniecības un rūpniecības ūdeņu ieplūšana Slocenē, kas ietek ezerā. Pēdējo divdesmit gadu laikā ezerā nogulsnējies un organismos uzkrājies apmēram 1000 t slāpekļa un vairāk nekā 100 t fosfora. Tas veicina ezera eitrofikāciju. Piesārņojuma ietekmē izzūd mieturaļģes (Chara) – nozīmīgs barības avots gan ezera bezmugurkauliniekiem, gan zivīm, gan arī putniem.
Nopietna problēma ir aizsardzības režīma neievērošana, malu zvejniecība (Smaļinskis, 1997).
3. Kaņiera ezera Cladocera fauna
2001.gada vasarā ![]()
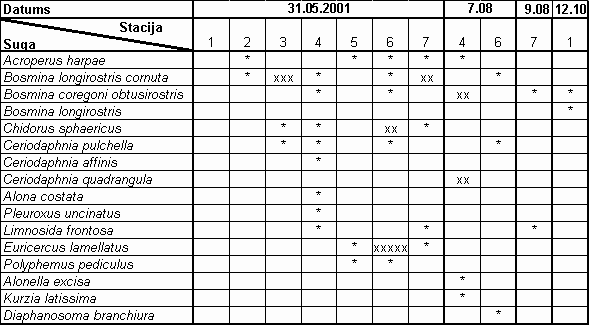
Kaņiera ezerā ir konstatētas 16 Cladocera sugas (1.tab), ko var uzlūkot par bagātu faunu eitrofajam ezeram (3.paraugā (31.05.2001) dominē Bosmina longirostris cornuta eitrofā forma). Lielā daudzveidība var tikt izskaidrota ar biezu zemūdens un piekrastes augāju. Sakarā ar ezera zivju krājumu ikgadēju palielināšanu, dažas sugas, iespējams, ir ievazātas.
Vislielākais sugu skaits (8 sugas) ir konstatētas 4. stacijā (31.05.2001), kas atrodas tuvu niedru audzēm, pie tam 3 no 8 sugām (Ceriodaphnia affinis, Alona costata, Pleuroxus uncinatus) netika atrastas nevienā citā paraugā ieskaitot vēlākā laikā ievākto šīs vietas paraugu.
Purvainajā vietā ar šaurlapu vilkvālītes audzēm ievāktajā 1. paraugā (31.05.2001) Cladoceranetika konstatētas, paraugā pārsvarā detrīts. No tā var secināt, ka 1. stacijas vieta nav piemērota Cladocerafaunai.
Sugu sastāvs 5., 6. un 7. stacijās ievāktajos paraugos (31.05.2001) ir ļoti līdzīgs, taču 6. stacijā zooplanktona daudzums ir ievērojami lielāks, kas liecina par šīs atklātās ūdens platības piemērotību kladocerām.
Vēlākajā laikā ievāktajos paraugos (7.8.2001 un 9.08.2001) ir konstatētas visumā 9 sugas, no kurām 4 (Ceriodadhnia quadrangula, Alonella excisa, Kurzia latissima, Diaphanosoma brachiurum) parādās tikai šā laika paraugos. Savukārt, Bosmina longirostris ir atrodama tikai oktobra (12.10.2001) paraugā.
Jaunu sugu parādīšanās liecina par zināmu sezonalitāti Cladocera ezera faunā, kurai netiek pakļautas visas sugas –Acroperus harpae, Bosmina longirostris cornuta, Ceriodaphnia pulchella, Limnosida frontosa un īpaši Bosmina coregoni obtusirostris ir sastopamas no pavasara līdz vasaras beigām.
1.tabula
Kaņiera ezera Cladocera fauna
Paraugu raksturojums:
1 (31.05) – totāli smalks augu detrīts, Cladocera nav;
2 (31.05) –lielāki detrīta sakopojumi, kaļķa inkrustācijas, iespējams, sadalījušas kladoceru čaulas. Pārsvarā – Rotatoria un Copepoda naupliji.
3 (31.05) – daudz liela kūdraina un aļģu detrīta, daudz zooplanktona atlūzas, bagāts zooplanktons (Rotatoria, Copepoda);
4 (31.05) – paraugā smiltis, augu detrīts. Dominē Bosmina;
5 (31.05) – smalks kūdrains detrīts, nedaudz fitoplanktona, zooplanktona maz;
6 (31.05) – daudz lielāka izmēra detrīta;
7 (31.05) – ļoti bagāts zooplanktons, masveidā Rotatoria;
6 (7.08) – detrīta nav, nedaudz aļģu koloniju, bagātīgi Copepoda un Rotatoria;
7 (9.08) – Daudz naupliju, arī Rotatoria;
1 (12.10) – ļoti maz aļģu, detrīta, zooplanktona;
1. Bērziņš B. 1966. Planktontab.III, tab.9-15
2. Brönmark Ch., Hansson L.-A. 2001. The biology of lakes and ponds. Oxford: Oxford, 216 pp.
3. Crease T., Taylor D. 1998. The origin and evolution of variable-region helices in V4 and V7 of the small subunitribosomal RNA of branchiopod crustaceans.- Molecular Biology and Evolution, 15: 1430-1446.
4. Dodson S., Frey D. 1991. Cladocera and other Branchiopoda.- Ecology and Classification of Notth American invertebrates, 723- 786.
5. Horne A., Cordman Ch. 1994. Limnology. International editions. 576 pp.
6. Lūmane H. 1995. Kaņiera ezers.- Latvijas daba, 2. sēj. 233 lpp.
7. Sloka N. 1961. Materiāli par Vidzemes Centrālās augstienes lielāko ezeru – Alauksta, Ineša, Kāla, Kaķiša – limnoekoloģiju.- P.Stučkas LU zinātniskie raksti, 39.sēj. 153-182 lpp.
8. Sloka N. 1971. Bezmugurkaulnieku zooloģijas praktikums. Rīga: Zinātne. 266 lpp.
9. Sloka N. 1981. Latvijas PSRS dzīvnieku noteicējs. Latvijas kladoceru (Cladocera) fauna un noteicējs. Rīga: P.Stučkas LU, 145 lpp.
10. Smaļinskis J. 1997. Der Kaņieris See. Rīga: Jāņa sēta, EUCC
11. Wetzel R. 1995. Limnology, Inland aquatic ecosystems. -Encyclopedia of Environmental biology. Vol.2 433-441 pp.
12. Zandmane A. 1995. Praktiskās hidrobioloģijas rokasgrāmata. Upju piesārņojuma bioloģiskās analīzes metodes. Rīga: LU ekoloģiskā centra apgāds “Vide”. 70 lpp.
13.Большая Советская Энциклопедия. 1975. Т.19 Москва: «Советская Энциклопедия » 633 стр.
14. Догель В. 1974. Зоология безпозвоночных. Москва: Высшая школа. 559 стр.
15. Константинов А. 1986. Общая гидробиология. Москва: Высшая школа. 472 стр.
16. Мануйлова Е. 1964. Ветвистоусые рачки (Cladocera) фауны СССР. Москва: Наука. 327 стр.
17.Яшнов В. 1969. Практикум по гидробиологии. Москва: Высшая школа. 428 стр.
18. www.ezeri.lv
19.www.jurmala.lv
20.www.kc-denmark.dk/public_html/p26.htm
21.www.valmiera.lanet.lv